

Cellemembraner er tynne hinner som omslutter alle celler. Cellemembranen skiller cellens innhold (cytoplasma) fra det ytre (ekstracellulære) miljøet. Cellemembranen kalles også plasmamembran.
Faktaboks
- Uttale
-
s'ellemembran
- Også kjent som
-
biologiske membraner
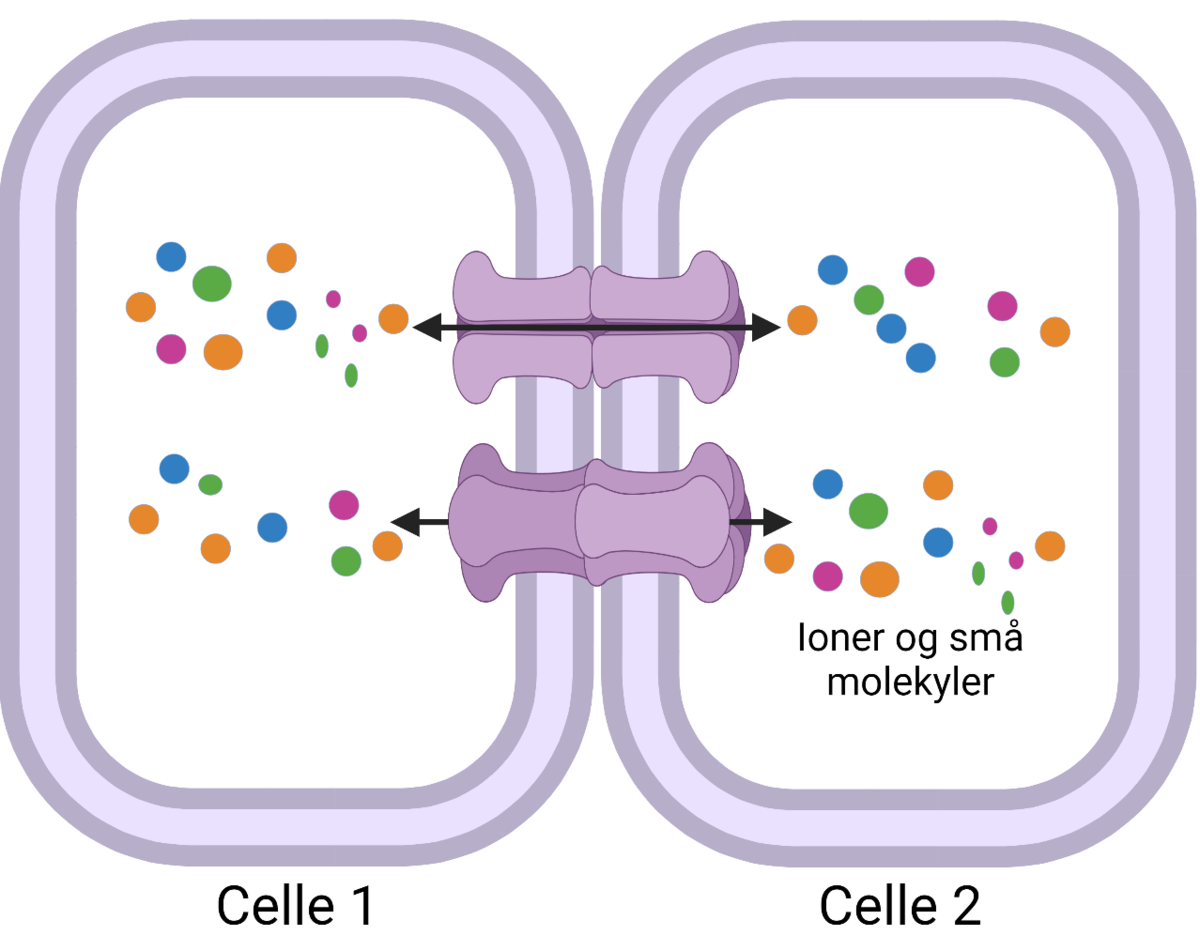
Cellemembranen har flere viktige oppgaver i tillegg til å omslutte cellen; den kontrollerer alt som kommer inn og ut av cellen og den regulerer hvordan celler kommuniserer både med andre celler og det ekstracellulære miljøet.
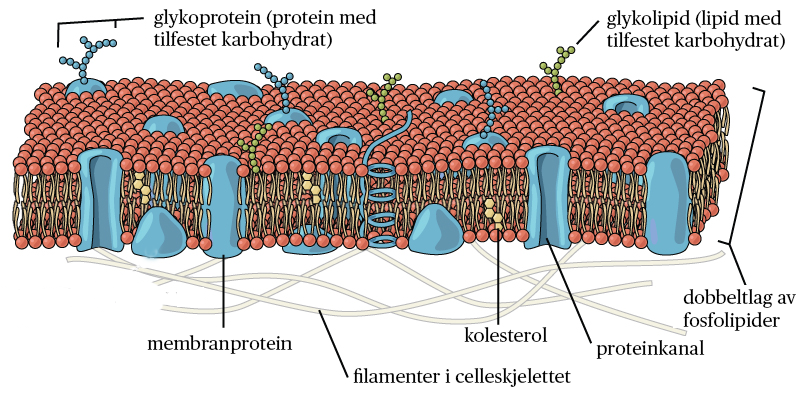
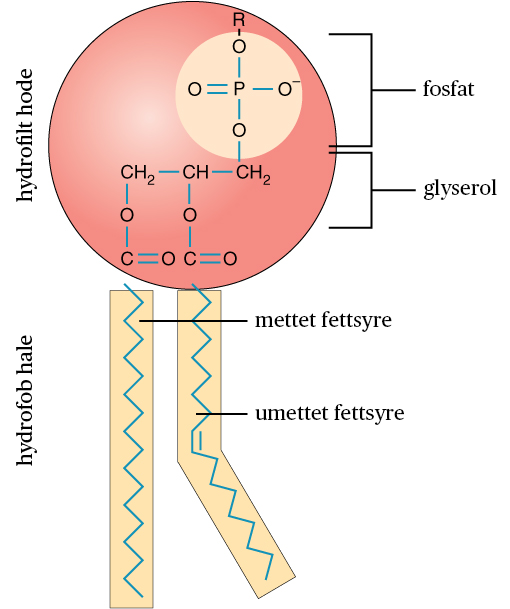
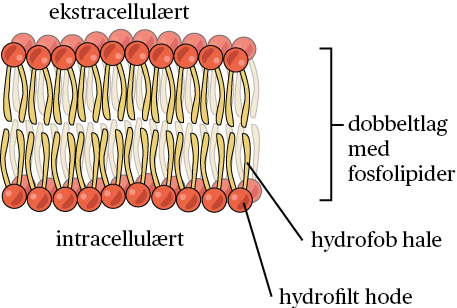
Cellemembranen består hovedsakelig av fettstoffer i form av fosfolipider som består av to fettsyrekjeder («haler») koplet til et «hode» av glycerol og fosfat. Fosfolipidene danner spontant et dobbeltlag slik at «hodene» peker ut av dobbeltlaget, mens «halene» peker inn i dobbeltlaget.
Mellom fosfolipidhodene flyter det proteiner som har ulike oppgaver. Noen går helt gjennom fosfolipidlaget, andre ligger delvis inn i laget.
Andre membraner i cellen, som dem som omslutter organeller, har liknende struktur. Alle membraner går ofte under samlebetegnelsen biologiske membraner.




Kommentarer (2)
skrev Milla Refsnes-Jensen
Fosfolipidene er amfipatiske molekyler med et hydrofilt (vannelskende) hode og en hydrofob (vannhatende) hale. Det hydrofobe hodet består av glycerol og fosfat, mens den hydrofile halen består av to fettsyrekjeder. Fettsyrekjedene kan være mettet eller umettet.
Du har blandet om hydrofob og hydrofil i siste setning der og når jeg søkte på google kom det opp og forvirret meg veldig.
svarte May-Britt Five
Takk for kommentar, det er rettet opp i nå.
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.