
Bivalvia
| Bivalvos | ||
|---|---|---|
| Rango temporal: Cámbrico-Holoceno | ||
 Bivalvos | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Mollusca | |
| Clase: |
Bivalvia Linnaeus, 1758 | |
| Subclases | ||
Subclase Anomalosdesmata Subclase Heterodonta Subclase Palaeoheterodonta
Subclase Protobranchia Subclase Pteriomorphia | ||
Los bivalvos (Bivalvia; bi: ‘dos’, valvia: ‘valva’ o ‘placa’), lamelibranquios (Lamellibranchia) o pelecípodos (Pelecypoda) son seres vivos clasificados biológicamente en el filo (tipo de organización) Mollusca (de molusco). Existen aproximadamente unas 13 000 especies de moluscos bivalvos, la mayoría marinas.[cita requerida] Presentan un caparazón con dos valvas laterales, que se cierran por acción de uno o dos músculos aductores. Estas son simétricas, generalmente, unidas por una bisagra y ligamentos.
Se les encuentra enterrados en fondos blandos (infauna), como habitantes fijos de superficies y estructuras rígidas o libres sobre los fondos (epifauna). Algunas especies perforan el sustrato (roca o madera) y algunas más son comensales o parásitas.[cita requerida]
Características
[editar]Carecen de cabeza diferenciada y de tentáculos. No tienen maxilas ni rádula.[1][2] La boca presenta palpos labiales carnosos. Respiran mediante uno o dos pares de branquias, generalmente laminares. Son invertebrados, es decir que no tienen huesos.
En las conchas de los bivalvos se observa gran variedad de tamaños, formas, colores y dibujos esculpidos en la superficie. El tamaño fluctúa desde conchas diminutas (2 mm) hasta especies que pueden alcanzar 15 dm de largo y un peso de 250 kg.[cita requerida] Entre los moluscos bivalvos más conocidos podemos nombrar: ostra, almeja, navaja, mejillón, broma de los barcos, coquina, etc.
En el borde anterior del manto se distinguen tres pliegues: interno, medio y externo. El pliegue interno es muscular, el medio destaca por su función sensorial y el externo está relacionado con la secreción de la concha. La concha está dividida en dos valvas unidas dorsalmente por la charnela, que consiste en un ligamento elástico formado por conquiolina y secretado por el manto; no está muy calcificado, por lo que permanece flexible y elástico. El músculo o "pie" característico de los moluscos, en los bivalvos puede presentarse modificado o muy reducido según el hábito de las diferentes especies.
-
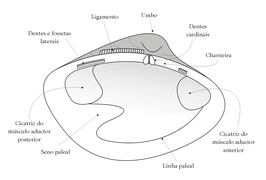 Vista interna de una valva.
Vista interna de una valva. -
 1: Plano de simetría. 2: Líneas de crecimiento. 3: Ligamento. 4: Umbo.
1: Plano de simetría. 2: Líneas de crecimiento. 3: Ligamento. 4: Umbo.


Músculos y ligamentos
[editar]El principal sistema muscular de los bivalvos son los músculos aductores posterior y anterior. Estos músculos conectan las dos valvas y se contraen para cerrar la concha. Las valvas también están unidas dorsalmente por el ligamento bisagra, que es una prolongación del periostraco. Este ligamento es responsable de la apertura del caparazón y actúa contra los músculos aductores cuando el animal se abre y se cierra.[3] Los músculos retractores conectan el manto al borde del caparazón, a lo largo de una línea conocida como línea palial.[4][5] Estos músculos tiran del manto a través de las válvulas..[5]
En los bivalvos sedentarios o yacentes que se apoyan sobre una sola valva, como las ostras y las vieiras, el músculo aductor anterior se ha perdido y el músculo posterior se sitúa en posición central.[6] En las especies que pueden nadar batiendo sus valvas, existe un único músculo aductor central. Estos músculos se componen de dos tipos de fibras musculares, haces musculares estriados para acciones rápidas y haces musculares lisos para mantener una tracción constante.[7] Los músculos protractor y retractor pedales emparejados accionan el pie del animal. [8][9][10]
Sistema nervioso
[editar]Los hábitos sedentarios de los bivalvos han hecho que, en general, el sistema nervioso sea menos complejo que en la mayoría de los demás moluscos. Los animales no tienen cerebro; el sistema nervioso consiste en una red nerviosa y una serie de ganglios emparejados. En todos los bivalvos, salvo en los más primitivos, hay dos ganglios cerebropleurales a cada lado del esófago. Los ganglios cerebrales controlan los órganos sensoriales, mientras que los ganglios pleurales suministran nervios a la cavidad del manto. Los ganglios pedales, que controlan el pie, están en su base, y los ganglios viscerales, que pueden ser bastante grandes en los bivalvos nadadores, están bajo el músculo aductor posterior. Ambos ganglios están conectados a los ganglios cerebropleurales por fibras nerviosas. Los bivalvos con sifones largos también pueden tener ganglios sifonales para controlarlos.[11][12]
Sentidos
[editar]Los órganos sensoriales de los bivalvos están situados en su mayor parte en los márgenes posteriores del manto. Suelen ser mecanorreceptores o quimiorreceptores, en algunos casos situados en tentáculos cortos. El osfradio es un parche de células sensoriales situado bajo el músculo aductor posterior que puede servir para degustar el agua o medir su turbidez. Los estatocistos ayudan a los bivalvos a sentir y corregir su orientación.[12] En el orden Anomalodesmata, el sifón inhalante está rodeado de tentáculos sensibles a la vibración para detectar presas.[13] Muchos bivalvos carecen de ojos, pero algunos miembros de Arcoidea, Limopsoidea, Mytiloidea, Anomioidea, Ostreoidea y Limoidea tienen ojos simples en el margen del manto. Estos consisten en una fosa de células fotosensoriales y una lente[30] Las vieiras tienen ojos más complejos con una lente, una retina de dos capas y un espejo cóncavo.[14] Todos los bivalvos tienen células sensibles a la luz que pueden detectar una sombra que caiga sobre el animal.[11]
Circulación y respiración
[editar]
Los bivalvos tienen un sistema circulatorio abierto que baña los órganos con sangre (hemolinfa). El corazón tiene tres cámaras: dos aurículas que reciben la sangre de las branquias y un ventrículo único. El ventrículo es muscular y bombea la hemolinfa hacia la aorta y, de ahí, al resto del cuerpo. Algunos bivalvos tienen una sola aorta, pero la mayoría también tienen una segunda, normalmente más pequeña, que sirve a las partes posteriores del animal.[15] La hemolinfa normalmente carece de pigmento respiratorio.[16] En el género carnívoro Poromya, la hemolinfa tiene amebocitos rojos que contienen un pigmento de hemoglobina.[17]
Las branquias emparejadas están situadas en la parte posterior y consisten en filamentos tubulares huecos con paredes finas para el intercambio de gases. La demanda respiratoria de los bivalvos es baja, debido a su relativa inactividad. Algunas especies de agua dulce, cuando se exponen al aire, pueden abrir ligeramente la concha y se puede producir el intercambio de gases.[18][19] Las ostras, incluida la ostra del Pacífico (Magallana gigas), tienen respuestas metabólicas variables al estrés ambiental, observándose con frecuencia cambios en la tasa de respiración.[20]
Aparato digestivo
[editar]Modos de alimentación
[editar]La mayoría de los bivalvos se alimentan por filtración, utilizando sus branquias para capturar alimento particulado del agua, como el fitoplancton. Los protobranquios se alimentan de un modo diferente, raspando detritus del fondo marino, y éste puede ser el modo original de alimentación utilizado por todos los bivalvos antes de que las branquias se adaptaran a la alimentación por filtración. Estos bivalvos primitivos se aferran al fondo con un par de tentáculos en el borde de la boca, cada uno de los cuales tiene una sola palpa o aleta. Los tentáculos están cubiertos de mucosidad, que atrapa el alimento, y de cilios, que transportan las partículas hasta los palpos. Éstos clasifican las partículas, rechazando las inadecuadas o demasiado grandes para digerirlas y transportando las demás a la boca.[21]
En los bivalvos más avanzados, el agua se introduce en la concha desde la superficie ventral posterior del animal, pasa hacia arriba a través de las branquias y se duplica para ser expulsada justo por encima de la entrada. Puede haber dos sifones alargados y retráctiles que llegan hasta el lecho marino, uno para la corriente de agua inhalante y otro para la exhalante. Las branquias de los bivalvos filtradores se conocen como ctenidios y se han modificado mucho para aumentar su capacidad de capturar alimento. Por ejemplo, los cilios de las branquias, que originalmente servían para eliminar sedimentos no deseados, se han adaptado para capturar partículas de alimento y transportarlas en un flujo constante de moco hasta la boca. Los filamentos de las branquias también son mucho más largos que los de los bivalvos más primitivos, y se pliegan para crear un surco a través del cual se puede transportar el alimento. La estructura de las branquias varía considerablemente y puede servir para clasificar a los bivalvos en grupos.[21][22]
Unos pocos bivalvos, como la poromya granulata (Poromya granulata), son carnívoros y se alimentan de presas mucho más grandes que las diminutas microalgas que consumen otros bivalvos. Los músculos aspiran el agua a través del sifón inhalante, que se modifica en un órgano con forma de vaca, succionando a las presas. El sifón puede retraerse rápidamente e invertirse, poniendo la presa al alcance de la boca. El intestino está modificado para poder digerir grandes partículas de alimento.[17]
El género inusual, Entovalva nhatrangensis, es endosimbiótico y sólo se encuentra en el esófago de los pepinos de mar (holothuroidea). Tiene pliegues en el manto que rodean completamente sus pequeñas válvulas. Cuando el pepino de mar aspira sedimentos, el bivalvo deja que el agua pase por encima de sus branquias y extrae finas partículas orgánicas. Para evitar ser arrastrado, el pepino de mar se sujeta con hilos bisoños a la garganta del huésped y permanece sin daños.[23]
Aparato digestivo
[editar]El tubo digestivo de los bivalvos típicos consta de esófago, estómago e intestino. Los estómagos protobranquiales tienen un mero saco adosado, mientras que los bivalvos que se alimentan por filtración tienen una varilla alargada de moco solidificado denominada "estilo cristalino" que se proyecta hacia el estómago desde un saco asociado. Los cilios del saco hacen que el estilo gire, enrollando una corriente de moco que contiene alimento procedente de la boca y agitando el contenido del estómago. Este movimiento constante impulsa las partículas de alimento hacia una región de clasificación situada en la parte posterior del estómago, que distribuye las partículas más pequeñas hacia las glándulas digestivas y las más pesadas hacia el intestino.[24] El material de desecho se consolida en el recto y se expulsa en forma de gránulos a la corriente de agua exhalada a través de un poro anal. La alimentación y la digestión están sincronizadas con los ciclos diurnos y de las mareas.[25]
Los bivalvos carnívoros suelen tener estilos cristalinos reducidos y el estómago tiene paredes gruesas y musculosas, extensos revestimientos cuticulares y zonas de clasificación y secciones de la cámara gástrica disminuidas.[26]
Sistema excretor
[editar]Los órganos excretores de los bivalvos son un par de nefridios. Cada uno de ellos consta de un largo tubo glandular en forma de bucle, que se abre en el pericardio, y una vejiga para almacenar la orina. También tienen glándulas pericárdicas que recubren las aurículas del corazón o se adhieren al pericardio, y sirven como órganos de filtración adicionales. Los desechos metabólicos se expulsan de las vejigas a través de un nefridioporo cerca de la parte delantera de la parte superior de la cavidad del manto y se excretan.[27][28]
Reproducción y desarrollo
[editar]Los sexos suelen estar separados en los bivalvos, pero se conoce algún hermafroditismo. Las gónadas se abren en los nefridios o a través de un poro separado en una cámara sobre las branquias.[29][30] Las gónadas maduras de machos y hembras liberan esperma y huevos en la columna de agua. El desove puede ser continuo o estar provocado por factores ambientales como la duración del día, la temperatura del agua o la presencia de espermatozoides en el agua. Algunas especies "desovan por goteo", liberando gametos durante un periodo prolongado que puede extenderse durante semanas. Otras desovan en masa y liberan sus gametos por lotes o todos a la vez.[31]
La fecundación suele ser externa. Normalmente, se produce una breve fase que dura unas horas o días antes de que los huevos eclosionen en larvas trocóforas. Estas se desarrollan posteriormente en otro estado larvario y se establecen en el fondo marino y sufren metamorfosis para convertirse en adultos.[29][32] En algunas especies, como las del género Lasaea, las hembras aspiran agua con esperma a través de sus sifones inhalantes y la fecundación tiene lugar en el interior de la hembra. A continuación, estas especies crían a las crías dentro de la cavidad de su manto, liberándolas finalmente en la columna de agua como larvas velígeras o como juveniles reptantes.[33]
La mayoría de las larvas de bivalvos que nacen de huevos en la columna de agua se alimentan de diatomeas u otro fitoplancton. En las regiones templadas, alrededor del 25% de las especies son lecitotróficas y dependen de los nutrientes almacenados en la yema del huevo, donde la principal fuente de energía son los lípidos. Cuanto más largo es el periodo que transcurre antes de que la larva se alimente por primera vez, más grandes tienen que ser el huevo y la yema. El coste reproductivo de producir estos huevos ricos en energía es elevado y suelen ser más pequeños en número. Por ejemplo, la tellina del Báltico (Macoma balthica) produce pocos huevos de alto valor energético. Las larvas que eclosionan de ellos dependen de las reservas energéticas y no se alimentan. Al cabo de unos cuatro días, se convierten en larvas de estadio D, cuando desarrollan por primera vez válvulas articuladas en forma de D. Estas larvas tienen un potencial de dispersión relativamente pequeño antes de asentarse. El mejillón común (Mytilus edulis) produce 10 veces más huevos que eclosionan en larvas y pronto necesitan alimentarse para sobrevivir y crecer. Pueden dispersarse más ampliamente, ya que permanecen planctónicos durante mucho más tiempo.[34]
Los bivalvos de agua dulce tienen un ciclo vital diferente. El esperma se introduce en las branquias de la hembra con el agua inhalada y se produce la fecundación interna. Los huevos eclosionan en larvas gloquidias que se desarrollan dentro de la concha de la hembra. Más tarde se liberan y se adhieren parasitariamente a las branquias o aletas de un pez huésped. Al cabo de varias semanas se desprenden del hospedador, sufren una metamorfosis y se convierten en adultos sobre el sustrato.[29]
Algunas especies de la familia de los mejillones de agua dulce Unionidae, conocidos comúnmente como mejillones de bolsillo, han desarrollado una estrategia reproductiva inusual. El manto de la hembra sobresale de la concha y se convierte en una imitación de pez pequeño, con marcas parecidas a las de los peces y ojos falsos. Este señuelo se mueve en la corriente y atrae la atención de los peces reales. Algunos ven el señuelo como una presa, mientras que otros ven un congénere. Se acercan y el mejillón libera un gran número de larvas de sus branquias, rociando a los curiosos peces con sus pequeñas crías parásitas. Estas larvas de gloquidios se introducen en las branquias del pez, donde se adhieren y desencadenan una respuesta tisular que forma un pequeño quiste alrededor de cada larva. Las larvas se alimentan descomponiendo y digiriendo el tejido del pez dentro de los quistes. Al cabo de unas semanas se liberan de los quistes y caen al lecho del arroyo como moluscos juveniles.[35]
Historia natural
[editar]Todos los representantes de esta clase son acuáticos, tanto marinos como dulceacuícolas, y pueden encontrarse desde los límites superiores de la pleamar hasta las zonas abisales. La protección de las conchas permite que algunas especies especializadas soporten las condiciones de la franja costera intermareal.
Típicamente, las especies que viven sobre sustratos blandos como fangos y arenas, presentan un pie que les permite excavar y tiene forma de hacha. Las especies sésiles se mantienen adheridas al sustrato, bien sea por cimentación, como las ostras, o mediante la secreción de una serie de filamentos que conforman el biso (mejillones).
Aquellos bivalvos que viven bajo la arena (suelo blando), se alimentan filtrando pequeñas cantidades de agua durante las mareas altas, de donde extraen el alimento. No suelen viajar lejos, ya que sus órganos están adaptados para filtrar el agua a través de las valvas de su concha en vez de que el bivalvo mismo se traslade para introducir agua en su interior. En las ocasiones en que se mueve, utiliza su pie, un músculo que le permite enterrarse en la arena.
Ecología y paleontología
[editar]
Los bivalvia son moluscos esencialmente acuáticos, en su mayoría marinos, que habitan con preferencia la región bentónica sublitoral, aunque también se los encuentra hasta las profundidades abisales. La salinidad y la temperatura se cuentan entre los factores que más influyen en su distribución. Tienen gran valor en los análisis paleoambientales debido a su estrecha relación con el tipo de sustrato, la buena presencia en el registro fósil desde el Cámbrico, su gran diversidad y abundancia en variados paleoambientes, y excelente representación en los medios acuáticos actuales. Estos moluscos viven en un amplio rango de temperaturas del agua; en general el tamaño y espesor de la conchilla decrecen al disminuir la temperatura. Son más comunes en aguas bien oxigenadas, aunque algunas especies oportunistas o con adaptaciones especiales pueden vivir en ambientes pobres en oxígeno o incluso completamente anóxicos. Es amplio también el rango de salinidad que soportan, desde aguas dulces continentales hasta mares hipersalinos, pero son mucho más diversos en condiciones de salinidad marina normal.
La salinidad y la temperatura tienen poca influencia sobre los hábitos de vida, en cambio la depredación condiciona notablemente algunos aspectos generales de la morfología funcional de los bivalvos, como la secreción del biso o la excavación, aunque se conoce menos su influencia en el desarrollo de variantes morfológicas específicas para protegerse de depredadores (cementación, espinas sifonales). Los factores más importantes que afectan el modo de vida de los bivalvos son la disponibilidad de alimento, el tipo de sustrato y el movimiento del agua. La morfología funcional de las valvas está muy relacionada con el carácter del sustrato. En cuanto a la turbulencia del agua, puede decirse que en general prefieren ambientes con movimiento moderado del agua; en condiciones turbulentas predominan los cementantes, nidificadores, perforantes y excavadores rápidos.
El estudio de la diversidad morfológica de los bivalvos en relación con distintas estrategias de modo de vida ha demostrado que existen morfologías adaptativas recurrentes,[36] tipificadas cada una por conjuntos de caracteres específicos. Muchos bivalvos (especialmente los infaunales) viven con su comisura orientada en posición perpendicualr a la interfase entre el sustrato y el agua, pero existen algunos (comunes entre los epifaunales, aunque también los hay infaunales) que lo hacen con una de las valvas contra el sustrato o de manera que la comisura queda aproximadamente paralela u oblicua a la interfase. Estos últimos bivalvos se denominan pleurotéticos, y se reconocen por ser inequivalvos en algún grado (valvas derecha e izquierda con convexidad, espesor, ornamentación o aun coloración diferentes), pueden poseer valvas torsionadas, cuya comisura no se halla en un plano y, en el caso de los pleurotéticos epifaunales, son casi siempre monomiarios. Los pleurotéticos incluyen a todos los bivalvos cementantes, la mayoría de los apoyados, muchos bisados y algunos excavadores.
Sin embargo estos hábitos no son ejercidos con exclusividad, presentándose frecuentes combinaciones de los mismos. Algunas especies excavadoras (especialmente árcidos) emplean un débil biso para aumentar la estabilidad en los estratos blandos. Otras especies que viven fijas por el biso, pertenecientes a los géneros Isognomun y Barbatia, habitan introducidas a la manera de una cuña entre colonias de corales o rocas, igual que algunas especies nidificadoras libres. Muchas formas nadadoras, como los pectínidos, pasan buena parte de su existencia apoyadas en el fondo o fijas por el biso. Petricola pholadiformis normalmente vive como un excavador en sedimentos blandos o perforador en sustratos duros. En el canal de Beagle, Hiatella sólida se fija por el biso a los sustratos rocosos y al cachiyuyo, en latitudes menores es perforadora en arenas duras o incrustante de bivalvos y gastrópodos, y en Brasil vive fija sobre rocas, estrellas de mar y briozoos o entre tubos de poliquetos.[37]
Estas variaciones en hábito pueden producirse a lo largo de la ontogenia, como se deduce a veces de cambios morfológicos registrados en la conchilla.
La diversidad de formas que presentan las conchillas se correlaciona con la diversidad de hábitats ocupados y la variedad de modos de vida adoptados, y así grupos de bivalvos poco relacionados filogenéticamente pueden desarrollar conchillas con caracteres similares.[38]
Véase también
[editar]Referencias
[editar]- ↑ Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology, 7th edition. Cengage Learning. pp. 367-368. ISBN 978-81-315-0104-7.
- ↑ Dorit, Robert L.; Walker, Warren F. Jr.; Barnes, Robert D. (1991). Zoology. Saunders College Publishing. p. 672. ISBN 978-0-03-030504-7. (requiere registro).
- ↑ Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology, 7th edition. Cengage Learning. p. 369. ISBN 978-81-315-0104-7.
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. p. 21. ISBN 0-85238-234-0.
- ↑ a b Morton, Brian. «Bivalve: The mantle and musculature». Encyclopædia Britannica. Consultado el 5 de mayo de 2012.
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. pp. 5-6. ISBN 0-85238-234-0.
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. p. 18. ISBN 0-85238-234-0.
- ↑ Wells, Roger M. (1998). «Class Bivalvia». Invertebrate Paleontology Tutorial. State University of New York College at Cortland. Archivado desde el original el 28 de febrero de 2010. Consultado el 11 de abril de 2012.
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. pp. 28-30. ISBN 0-85238-234-0.
- ↑ Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology, 7th edition. Cengage Learning. pp. 371-372. ISBN 978-81-315-0104-7.
- ↑ a b Cofrancesco, Alfred F. (2002). «Nervous System and Sense Organs in Bivalves». Zebra Mussel Research Program. Archivado desde el original el 15 de abril de 2012. Consultado el 5 de mayo de 2012.
- ↑ a b Morton, Brian. «Mollusk: The nervous system and organs of sensation». Encyclopædia Britannica. Consultado el 8 de julio de 2012.
- ↑ Allen, J. A.; Morgan, Rhona E. (1981). «The functional morphology of Atlantic deep water species of the families Cuspidariidae and Poromyidae (Bivalvia): an analysis of the evolution of the septibranch condition». Philosophical Transactions of the Royal Society B (en inglés) 294 (1073): 413-546. Bibcode:1981RSPTB.294..413A. doi:10.1098/rstb.1981.0117.
- ↑ Colicchia, G.; Waltner, C.; Hopf, M.; Wiesner, H. (2009). «The scallop's eye—a concave mirror in the context of biology». Physics Education 44 (2): 175-179. Bibcode:2009PhyEd..44..175C. S2CID 122110847. doi:10.1088/0031-9120/44/2/009.
- ↑ Dorit, Robert L.; Walker, Warren F. Jr.; Barnes, Robert D. (1991). Zoology. Saunders College Publishing. p. 659. ISBN 978-0-03-030504-7. (requiere registro).
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. p. 36. ISBN 0-85238-234-0.
- ↑ a b Vaughan, Burton (2008). «The Bivalve, Poromya granulata». Archerd Shell Collection. Archivado desde el original el 7 de abril de 2018. Consultado el 3 de abril de 2012.
- ↑ Morton, Brian. «Bivalve: The respiratory system». Encyclopædia Britannica. Consultado el 8 de julio de 2012.
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. p. 25. ISBN 0-85238-234-0.
- ↑ Ducker, James; Falkenberg, Laura J. (2020). «How the Pacific Oyster Responds to Ocean Acidification: Development and Application of a Meta-Analysis Based Adverse Outcome Pathway». Frontiers in Marine Science (en inglés) 7. ISSN 2296-7745. doi:10.3389/fmars.2020.597441.
- ↑ a b Dorit, Robert L.; Walker, Warren F. Jr.; Barnes, Robert D. (1991). Zoology. Saunders College Publishing. p. 675–678. ISBN 978-0-03-030504-7. (requiere registro).
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. p. 87. ISBN 0-85238-234-0.
- ↑ Lützen, J.; Berland, B.; Bristow, G. A. (2011). «Morphology of an endosymbiotic bivalve, Entovalva nhatrangensis (Bristow, Berland, Schander & Vo, 2010) (Galeommatoidea)». Molluscan Research 31 (2): 114-124.
- ↑ Dorit, Robert L.; Walker, Warren F. Jr.; Barnes, Robert D. (1991). Zoology. Saunders College Publishing. p. 678–679. ISBN 978-0-03-030504-7. (requiere registro).
- ↑ Morton, Brian. «Bivalve: The digestive system and nutrition». Encyclopædia Britannica. Consultado el 7 de mayo de 2012.
- ↑ Tëmkin, Ilya; Strong, Ellen E. (2013). «New insights on stomach anatomy of carnivorous bivalves». Journal of Molluscan Studies 79 (4): 332-339. doi:10.1093/mollus/eyt031.
- ↑ Morton, Brian. «Bivalve: The excretory system». Encyclopædia Britannica. Consultado el 7 de mayo de 2012.
- ↑ Gosling, Elizabeth (2004). Bivalve Molluscs: Biology, Ecology and Culture. Fishing News Books. p. 37. ISBN 0-85238-234-0.
- ↑ a b c Dorit, Robert L.; Walker, Warren F. Jr.; Barnes, Robert D. (1991). Zoology. Saunders College Publishing. p. 682. ISBN 978-0-03-030504-7. (requiere registro).
- ↑ Morton, Brian. «Bivalve: The reproductive system». Encyclopædia Britannica. Consultado el 7 de mayo de 2012.
- ↑ Helm, M. M.; Bourne, N.; Lovatelli, A. (2004). «Gonadal development and spawning». Hatchery culture of bivalves: a practical manual. FAO. Consultado el 8 de mayo de 2012.
- ↑ Morton, Brian. «Bivalve: Reproduction and life cycles». Encyclopædia Britannica. Consultado el 7 de mayo de 2012.
- ↑ Ó Foighil, D. (1989). «Planktotrophic larval development is associated with a restricted geographic range in Lasaea, a genus of brooding, hermaphroditic bivalves». Marine Biology 103 (3): 349-358. S2CID 189852952. doi:10.1007/BF00397269.
- ↑ Honkoop, P. J. C.; Van der Meer, J.; Beukema, J. J.; Kwast, D. (1999). «Reproductive investment in the intertidal bivalve Macoma balthica». Journal of Sea Research 41 (3): 203-212. Bibcode:1999JSR....41..203H. doi:10.1016/S1385-1101(98)00053-7.
- ↑ Piper, Ross (2007). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. Greenwood Press. pp. 224–225. ISBN 978-0-313-33922-6. (requiere registro).
- ↑ Stanley, Steven M. (1970). Relations of shell form to life habits in the Bivalvia (Mollusca). Geological Society Memoir.
- ↑ Gordillo, S. (1995). Moluscos australes: Una guía ilustrada. Zaguier y Urruty Publications, Buenos Aires. p. 115.
- ↑ Camacho, H. H., Damborenea, S. E. and del Río, C. J. 2008. Bivalvia, en Camacho, H. H. and Longobucco, M. I. (eds.) Los Invertebrados fósiles. pp. 387–440. Fundación de Historia Natural Félix de Azara, Universidad Maimónides. ISBN 978-987-22121-7-9.
- Jonathan Elphick, Jen Green, Berbara Taylor, Richard Walker (2001). Gran Enciclopedia de los Animales. Santiago de Chile: La Tercera.
Enlaces externos
[editar] Wikispecies tiene un artículo sobre Bivalvia.
Wikispecies tiene un artículo sobre Bivalvia. Wikimedia Commons alberga una categoría multimedia sobre Bivalvia.
Wikimedia Commons alberga una categoría multimedia sobre Bivalvia.- Bivalvia - Malacología - Página de la Sociedad Española de Malacología.
| Control de autoridades |
|
|---|