
Homo erectus
| Homo erectus | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Solo insanının (Homo erectus soloensis) kafatası çizimi | |||||||||||||||||||||||
| Biyolojik sınıflandırma | |||||||||||||||||||||||
| |||||||||||||||||||||||
| İkili adlandırma | |||||||||||||||||||||||
| †Homo erectus (Dubois, 1894) | |||||||||||||||||||||||
| Alt türler | |||||||||||||||||||||||
Liste
| |||||||||||||||||||||||
| Sinonimler | |||||||||||||||||||||||
| |||||||||||||||||||||||
Homo erectus (Latincede "dik insan" anlamına gelir), en eski kalıntıları 2 milyon yıl öncesine tarihlendirilen, Pleistosen'de yaşamış soyu tükenmiş bir arkaik insan türüdür.[1] Örnekleri Homo (insan) cinsinin ilk tanınabilen üyeleri arasındadır.
H. heidelbergensis ve H. antecessor gibi çeşitli insan türlerinin H. erectus'tan, Neandertaller, Denisova insanları ve modern insanların da H. heidelbergensis'ten evrimleştiği kabul ediliyor.[2] H. erectus, İber Yarımadası'ndan Cava adasına dek uzanan kıtasal dağılımıyla Avrasya'ya yayılan ilk insan atasıydı. H. floresiensis[3] ve Homo luzonensis türleri, H. erectus'un Asya topluluklarından türemiş olabilir.[4] Bilinen son Homo erectus, 117.000-108.000 yıl önce yaşamış Solo insanıdır.[5]
H. erectus, atalarından daha çağdaş olan yürüyüşü ve vücut oranlarıyla ayırt edilir. Ayrıca düz bir yüze, belirgin bir burna ve muhtemelen seyrek vücut kıllarına sahip ilk insan türüdür. Türün beyin boyutu atalarınınkinden yüksektir ancak topluluklara bağlı olarak büyük ölçüde değişir. İlk H. erectus topluluklarında beyin gelişiminin çocukluğun erken döneminde durmuş gibi görünmesi, yavruların doğumda büyük ölçüde kendi kendine yeterli olduğunu ve dolayısıyla yaşam boyunca bilişsel gelişimlerinin sınırlı olduğunu gösteriyor.
H. erectus bir tepe avcıydı:[6] fosilleri genellikle sığırlar veya filler gibi orta ila büyük hayvanların kalıntıları ile birlikte bulunmuştur. Bunun gibi kanıtlar, insan atalarında ilk kez yırtıcı davranışın ve eşgüdümlü avlanmanın geliştiğini gösterir. Aşölyen taş alet kültürünü başlatan H. erectus'un ateş kullanma,[7] eşgüdümlü takımlar hâlinde avcılık, toplayıcılık, yaralı veya hasta toplum üyelerine bakma ve bunların yanı sıra muhtemelen denizcilik ve sanat yapma becerisine sahip ilk insan atası olduğu tahmin edilmektedir.
H. erectus'un erkek ve dişilerinin kabaca aynı büyüklükte olduğu sanılmaktadır, yani türde eşeysel dimorfizm azalmış olabilir. Eşeysel dimorfizmin azalması, primatlarda genellikle tek eşliliğin göstergesidir. Bu azalmaya karşın ağırlıkları 40 ila 68 kg, boyları ise 146 ila 185 cm aralığında değişkenlik gösteriyordu. H. erectus'un yapısal olarak konuşma yeteneğine sahip olup olmadığı belli değildir ancak bir tür ön-dil kullanarak iletişim kurdukları varsayılır.
Keşif ve adlandırma
[değiştir | kaynağı değiştir]
Charles Darwin'in 1871 tarihli İnsanın Türeyişi kitabında ifade ettiği, insanlığın Afrika'da evrimleştiği görüşünün aksine, 19. yüzyılın sonlarında yaşamış pek çok doğa bilimci, Avrupa ile Amerika'nın ortasında yer aldığı ve dünya çapında en uygun dağılım yollarını sağladığı için Afrika'nın değil Asya'nın insanlığın doğum yeri olduğunu ileri sürdü (Asya'dan çıkış kuramı). Bunlardan biri olan Ernst Haeckel, ilk insanın, şimdiki Güneydoğu Asya'da, var olmadığı kanıtlanmış efsanevi "Lemurya" kıtasında yaşamış "Pithecanthropus alalus" ("konuşamayan maymun insan" anlamına gelir) adını verdiği bir türden evrimleştiğini savunuyordu.[8] "Lemurya"nın Hint Okyanusu'nun altına battığı sanılıyordu, dolayısıyla bunu kanıtlayacak hiçbir fosil bulunamadı. Yine de Haeckel'in varsayımı Hollandalı bilim insanı Eugène Dubois'e Hollanda Doğu Hint Adaları'na seyahat etme konusunda esin oldu. Hiçbir yönlendirilmiş keşif gezisi insan fosili bulmadığından (bilinen az sayıdaki fosilin tamamı tesadüfen keşfedilmişti) ve ekonomi Uzun Buhran nedeniyle güç durumdayken, Hollanda hükümeti Dubois'e fon sağlamayı reddetti. 1887'de Hollanda Doğu Hindistan Ordusu'na sağlık subayı olarak katıldı ve 1887'de Hint Adaları'nda boş zamanlarında insan evrimini açığa kavuşturacak "kayıp halkasını" aramak üzere bir görev almayı başardı.[8] Cava'da, Solo Nehri kıyısındaki Trinil bölgesinde 1891'de üst bölümü korunmuş bir kafatası ve 1892'de bir uyluk kemiği buldu; bu fosile (Cava insanı) 1893'te Pithecanthropus erectus ("dik maymun insan") adını verdi. Avrupa bilim camiasını dik yürüyen bir maymun insan bulduğuna ikna etmeye çalıştı ama başarısız oldu. O zamanlar çok az sayıda insan atası fosilinin keşfedildiği göz önüne alındığında, bulgularını insan dışı, kusurlu bir maymun olarak büyük ölçüde görmezden geldiler.[9]
Bu fosillerin önemi, 1927 yılında Kanadalı paleoantropolog Davidson Black'in Çin'in Pekin yakınlarındaki Zhoukoudian mağarasında "Sinanthropus pekinensis" (Pekin insanı) adını verdiği türün keşfine kadar anlaşılamayacaktı. Black, o zamandan bu yana dünyanın en verimli H. erectus bölgesi hâline gelmiş[10] bölgeyi kazmaya devam etmek için finansman sağlamak üzere Kuzey Amerika ve Avrupa'da lobicilik faaliyeti yürüttü.[11] Cava'ya olan ilginin devam etmesi, 1931'de Ngandong'da (Solo insanı), 1936'da Mojokerto'da (Cava insanı) ve 1937'de Sangiran'da (Cava insanı) daha fazla H. erectus fosili keşfine yol açtı. Sangiran bölgesinde en iyi korunmuş Cava insanı kafatasını ortaya çıkarıldı.[12] Alman paleoantropolog Franz Weidenreich, Çin örneklerinin ayrıntılı açıklamalarını çeşitli monografilerinde yaptı. Özgün örnekler, İkinci Çin-Japon Savaşı sırasında, korunmak üzere Çin'den kaçırılma girişiminin ardından kayboldu. Günümüze kalıntıların yalnızca alçı dökümleri kaldı.
_presented_at_Paleozoological_Museum_of_China.jpg)
Cava insanı ile Pekin insanı arasındaki benzerlikler, Ernst Mayr'ın 1950'de her ikisini de Homo erectus olarak yeniden adlandırmasına yol açtı. 20. yüzyılın büyük bir kısmı boyunca antropologlar H. erectus'un insan evrimindeki rolünü tartıştılar. Yüzyılın başlarında, kısmen Cava ve Zhoukoudian'daki keşifler nedeniyle, modern insanın ilk olarak Asya'da evrimleştiği inancı geniş çapta kabul gördü. Ancak birkaç doğa bilimci (en önde Charles Darwin) insanların en eski atalarının Afrikalı olduğu kuramını savundular. Darwin, insanın en yakın akrabaları olan şempanze ve gorillerin yalnızca Afrika'da evrimleştiğine ve yaşadığına dikkat çekmişti.[13] Darwin orangutanları Eski Dünya'nın büyük insansı maymunları arasına dahil etmemişti, bunun nedeni muhtemelen orangutanları insansı maymundan ziyade ilkel insanlar olarak düşünmesiydi.[14] Darwin, Afrika'yı insanın atalarının en muhtemel doğum yeri olarak değerlendirirken, İnsanın Türeyişi ve Cinsel Seçilim adlı kitabında insanın kökeninin coğrafi konumu hakkında da şu ifadeyi kullanmıştır: "(...) bu konu üzerinde kurgu yapmanın yararı yok; iki veya üç antropomorfik [insan biçimli] insansı maymun, biri Dryopithecus (...), Miyosen devresinde Avrupa'da vardı ve bu kadar uzak bir dönemden bu yana, dünya kesinlikle pek çok büyük değişimden geçti, devrimler oldu ve [Afrika'dan] en geniş ölçekte göç için yeterli zaman oluştu." (1889, ss. 155-156).
1949'da Güney Afrika'daki Swartkrans Mağarası'nda bulunan H. erectus kalıntıları paleoantropologlar Robert Broom ve John Talbot Robinson tarafından "Telanthropus capensis" adında bir tür olarak tanımlandı.[15] Yakındaki mağaralardan da Homo cinsine ait fosiller olduğu bildirildi ancak bunların türlerinin belirlenmesi çalkantılı bir tartışma oldu. Birkaç Kuzey Afrika bölgesinde daha H. erectus kalıntıları bulunmuş ve ilk olarak 1951'de "Atlanthropus mauritanicus" olarak sınıflandırılmıştır.[10] 1970'li yıllardan itibaren, özellikle Richard Leakey'in öncülüğünde, Doğu Afrika'da, ağırlıklı olarak Kenya'daki Koobi Fora bölgesinde ve Tanzanya'daki Olduvai Gorge'da daha fazla fosil gün yüzüne çıkarıldı.[10]
Avrupa genelinde ortaya çıkarılan arkaik insan fosilleri, önceleri H. erectus'a atfedilirken, İngiliz insan bilimci Chris Stringer'ın çalışmaları sonucunda H. heidelbergensis olarak ayrı bir türde sınıflandı.[16]
Evrim
[değiştir | kaynağı değiştir]
H. erectus'un yaklaşık 2 milyon yıl önce H. habilis'ten evrimleştiği öne sürülüyor ancak bu görüş, en az yarım milyon yıl boyunca bir arada var olduklarından sorgulanıyor. Başka bir görüşe göre, bir grup H. habilis üreme yoluyla yalıtılmış olabilir ve yalnızca bu grup H. erectus'a (kladogenez) evrimleşmiştir.[17]
H. erectus'un en eski kalıntıları hem Afrika'da hem de Doğu Asya'da bulunduğundan (Çin'de 2,1 milyon yıl kadar erken,[18][19][20] Güney Afrika'da 2,04 milyon yıl önce[1][21]), H. erectus'un nerede evrimleştiği tartış��lmaktadır. 2011 yılında yapılan bir araştırma, H. habilis'in Afrika'dan Batı Asya'ya ulaştığını, ilk H. erectus'un burada evrimleştiğini ve erken H. erectus'un daha sonra Batı Asya'dan Doğu Asya (Pekin insanı), Güneydoğu Asya (Cava insanı) ve Avrupa'da (Tautavel insanı) dağıldığı ayrıca bir grubun Afrika'ya geri döndüğü (Homo ergaster) ve modern insanların bu göçmenlerden türediği varsayımını ortaya attı.[22][23] Diğer insan bilimciler H. erectus/H. ergaster'in Afrika'da evrimleştiği ve sonunda burada modern insanlara dönüştüğünü savunuyor.[24][25]
H. erectus 1,8 milyon yıl önce Cava'nın Sangiran kentine ulaşmıştı[26] ve H. erectus'un ikinci ve farklı bir göç dalgası yaklaşık 780 bin yıl önce Çin'in Zhoukoudian kentine yerleşmişti. Sangiran'da bulunan H. erectus'ların dişleri daha büyüktür ve Zhoukoudian'daki H. erectus'un gelişmiş dişlerinden çok ilkel (atasal) Batılı H. erectus ve H. habilis'in dişlerine daha benzerdir. Ancak sonraki dönemlerde Sangiran H. erectus'larının diş boyutlarının küçüldüğü görülüyor. Bu, Cava'ya Zhoukoudian veya onunla akraba olan bazı topluluklar tarafından yapılmış ikincil bir göç olayına işaret ediyor olabilir.[27]

Torunları ve sinonimleri
[değiştir | kaynağı değiştir]Homo erectus, neredeyse iki milyon yıl boyunca hayatta kalan, Homo'nun en uzun ömürlü türüdür. Buna karşılık modern insan (Homo sapiens) 300.000 yıl kadar önce ortaya çıktı.
Pek çok arkaik insanın, H. erectus'un veya H. sapiens'in alt türü olarak mı yoksa ayrı türler olarak mı sınıflandırılması gerektiği konusunda kesin bir fikir birliği yoktur.
- Afrikalı H. erectus adayları
- Homo ergaster (veya "Afrikalı H. erectus")
- Homo naledi
- Avrasyalı H. erectus adayları:
- Homo floresiensis[28]
- Homo rhodesiensis
- 1982 yılında Hindistan'ın Madhya Pradesh kentinde bulunan Narmada fosili ilk başta H. erectus veya Homo erectus narmadensis olarak tanımlanmıştı.[29]
Cava'da bulunan ve 1,4 ila 0,9 milyon yıl öncesine tarihlenen fosillere dayanan Meganthropus, dev bir erken insan türü olduğu yönündeki daha önceki yorumların aksine[30] geçici olarak H. erectus türü altında gruplandırıldı ancak daha eski çalışmalar fosilleri Homo cinsiyle yakın akraba olmayan bir insansı maymun olarak tanımlamıştı.[31] Zanolli vd. (2019), Meganthropus'un soyu tükenmiş insansı maymunların ayrı bir cinsi olduğuna karar verdi.[32]
Anatomi
[değiştir | kaynağı değiştir]Baş
[değiştir | kaynağı değiştir]
Homo erectus diğer erken homininlere göre düz bir yüze sahipti; belirgin bir kaş sırtı, görece alçak ve düz bir yüzü vardı.[33][34] Kafatası eklemleri boyunca uzanan küçük tepeler olan sagital, frontal ve taç çıkıntılarının varlığının, kafatasının, özellikle de kafatası kubbesinin önemli ölçüde kalınlaştığının kanıtı olduğu öne sürülmüştür. Ancak bilgisayarlı tomografi taramaları durumun böyle olmadığını ortaya koyuyor. Bununla birlikte, kafatasının arka kısmındaki artkafa kemiğinin skuamöz kısmı, özellikle de iç artkafa tepesi, modern insanınkinden belirgin şekilde daha kalındır ve bu muhtemelen ilkel bir özelliktir.[34][35]
Fosil kayıtları, H. erectus'un, genellikle nemi korumak için kuru havayı solumaya yanıt olarak evrimleştiği düşünülen, çıkıntılı bir buruna sahip ilk insan türü olduğunu göstermektedir.[36] Amerikalı ruh bilimci Lucia Jacobs, çıkıntılı burnun, yol bulmayı ve uzun mesafeli göçü kolaylaştırmak için kokuların geldiği yönü (stereo koku alma) ayırt etmeye yaradığını ve bu yüzden evrimleştiğini önerdi.[37]
Asyalı H. erectus'un ortalama beyin büyüklüğü yaklaşık 1000 cm3'tür. Bununla birlikte, Gürcistan'ın Dmanisi kentinde (H. e. georgicus), Kenya'nın Koobi Fora ve Olorgesailie bölgelerinde ve Etiyopya'nın Gona bölgesinde beyin boyutu çok daha küçük kafatası örnekleri bulunmuştur. Genel olarak H. erectus'un beyin boyutu 546 ila 1251 cm3 arasında değişmektedir;[38] bu, modern insanlarda ve şempanzelerde görülen çeşitlilik aralığından çok daha büyüktür.

2021 yılında yayımlanan "İnsan beyni boyutunda topluluklar arası çeşitlilik: Hominin bilişsel filogenisine yönelik çıkarımlar" başlıklı bir makalede, Asyalı H. erectus'un son 600 bin yıldaki beyin boyutunun modern insan topluluklarıyla önemli ölçüde örtüştüğü belirlendi. Araştırmadaki önemli bir nokta, bazı küçük beyinli modern insan topluluklarının H. erectus'a, vücutları ve beyinleri büyük olan modern insan topluluklarına göre daha çok benzerlik göstermesiydi. Makale, tür ortalamalarının fosillerle karşılaştırıldığı ve topluluklar arası çeşitliliğin göz ardı edildiği, insan evrimindeki beyin büyüklüğü artışına ilişkin mevcut anlayıştaki yöntemsel kusurlara dikkat çekiyor. Aynı zamanda, bazı modern insan topluluklarının H. erectus'a göre beyin büyüklüğünde çarpıcı bir artış yaşamadığı, artışın çoğunun kuzey topluluklarında meydana geldiği ve bunun da topluluklar arası çeşitliliğin farkına varılamamasına neden olduğu gerçeğine de değiniyor. Yazarlar bu kusurların nereden kaynaklandığını şöyle anlatıyor: "... H. sapiens'in ortalama kafatası büyüklüğündeki artış, büyük ölçüde üst sınırdaki artıştan kaynaklanmaktadır ve bizim H. erectus örneğimize göre alt sınırdaki artış çok daha az belirgindir. Bunun yanında üst sınırdaki bu artışın kuzeydeki topluluklarda daha belirgin olduğu görülüyor; bu, iklim etkenlerine ek olarak vücut büyüklüğündeki göreli artışın bir sonucu olabilir." Sonuç olarak, daha önceki araştırmacıların da önerdiği gibi, tamamen beyin boyutu benzerliklerine dayanarak Asyalı H. erectus'un H. sapiens'in bir alt türü, yani H. sapiens soloensis olarak yeniden sınıflandırılabileceğini öne sürüyorlar.[39]
Diş açısından H. erectus, Plio-Pleyistosen homininleri (Australopithecus vs.) arasında en ince diş minesine sahiptir. Mine, dişin sert yiyecekler yüzünden kırılmasını engellemekle birlikte sert yiyecekleri kırmayı da engeller. H. erectus'un ve tüm erken insan (Homo) türlerinin alt çene kemiklerinin gövdeleri, modern insanların ve yaşayan tüm insansı maymunlarınkinden daha kalındır. Alt çene gövdesi, ısırma kuvveti veya çiğneme nedeniyle bükülmeye karşı direnç gösterir, bu da çenelerinin yemek yerken alışılmadık derecede güçlü gerilmeler üretebileceği anlamına gelir ancak bunun nasıl uygulandığı belirsizdir. Bununla birlikte H. erectus'un alt çene gövdeleri erken insan türlerinden biraz daha incedir. Ayrıca azı ve küçük azılarda H. habilis'e göre çukur sıklığı daha fazladır; bu da H. erectus'un daha kırılgan yiyecekler yediğini düşündürür. Bunların hepsi, H. erectus ağzının sert gıdaları ezme konusunda daha az, kesme konusunda ise daha yetenekli olduğunu, dolayısıyla muhtemelen alet kullanımına bir tepki olarak beslenebileceği gıda çeşitliliğini azalttığını gösteriyor.[40]
Vücut
[değiştir | kaynağı değiştir]
Modern insanlara benzer biçimde H. erectus'un boyu 146–185 cm ve ağırlığı 40–68 kg arasında değişiyordu. Bu çeşitliliğin iklim, ölüm oranları veya beslenmedeki bölgesel farklılıklardan kaynaklandığı düşünülüyor.[41][42] Primatlar arasında, çevresel zorluk etkenlerine verilen bu belirgin tepkinin (fenotipik esneklik) varlığı yalnızca modern insanlarda gözlemlenmiştir.[43][44]
Modern insanlara benzer ve diğer büyük insansı maymunlardan farklı olarak, H. erectus'un erkekleri ve dişileri arasında büyük bir boyut farklılığı (boyuta özgü eşeysel dimorfizm) yok gibi görünüyor. Ancak bunu doğrulayabilecek fosil verileri oldukça azdır.[45] Örneğin Koobi Fora'dan iki yetişkin örneğin beyin büyüklüğü 848 ve 804 cm3 olarak,[38] önemli ölçüde daha küçük olan başka bir yetişkin örneğin ise 691 cm3 olarak ölçüldü; bu, örneklerin cinsiyeti belirsiz olsa da eşeysel dimorfizme işaret edebilir.[17] Fosil kayıtlarında cinsiyeti belirlemenin zorluğunu gösteren bir diğer örnek ise Olduvai Vadisi'nde bulunan birkaç örnektir. 1960 yılında Olduvai Vadisi'nde bulunan ve OH12 ve OH9 olarak tanımlanan iki H. erectus kafatasının beyin boyutları 1000 cm3 ve 700 cm3 olarak hesaplanmıştır.[46] Eşeysel dimorfizmin belirli özellikleri, genellikle kas işaretinin olmaması gibi cinsiyeti belirleme olanaklarıyla tanımlanır[47] ve kalıntılar parçalı olduğundan burada eşeysel dimorfizmin söz konusu olup olmadığı belli değil.[46] Eğer H. erectus'ta eşeysel dimorfizm yoktuysa, muhtemelen H. erectus insan soyunda bu özelliğe sahip ilk türdür. Bunun için de o dönemde dişilerin boyutunda önemli ve ansızın bir artış olması gerekir. Daha önceki türlere ait parçalı fosiller, eşeysel dimorfizmdeki bu azalmanın ne zaman olduğunu belirsiz kılmaktadır.[48]

H. erectus'un modern insanlarla hemen hemen aynı üye (kol ve bacak) biçimlerine ve oranlarına sahip olması, modern insan benzeri bir hareket yeteneği olduğunu düşündürüyor.[49][50] Kenya'nın Ileret kenti yakınlarındaki H. erectus izleri de bu görüşü destekliyor.[51] Modern insandakine benzer omzu ise nesneleri hızlıca fırlatabilme yeteneğini akla getiriyor.[52] Bir zamanlar Turkana Çocuğu'nun modern insanlarda görülen 5 bel omuru yerine 6 omura sahip olduğu, göğüs omurlarının ise 12 yerine 11 tane olduğu düşünülüyordu ancak daha sonra bu düşünceden vazgeçildi. Artık örneğin insan benzeri bir omurga eğriliğine sahip olduğu ve omur sayılarının modern insanlarla aynı olduğu kabul ediliyor.[53]
İnsan atalarının vücut kıllarının çoğunu ne zaman kaybettiği büyük oranda belirsizdir. Genetik çözümleme, koyu tene neden olan melanokortin 1 reseptöründeki yüksek etkinliği 1,2 milyon yıl öncesine tarihliyor. Vücut kıllarının eksikliği cildi zararlı morötesi ışınıma maruz bırakacağından, tenin koyulaşması kılsızlaşmanın göstergesi olabilir.[54] Açıkta kalan deri ancak Pleistosen'de güneş ışığına uyumsuz hâle gelmiş olabilir çünkü Dünya'nın artan eksen eğikliği (ki bu aynı zamanda buzul çağlarına da neden olmuştur) güneş ışınımının yeryüzündeki etkisini artırmış olacaktır, bu da kılsızlığın ilk olarak australopitesinlerde ortaya çıktığını düşündürmektedir.[55] Bununla birlikte australopitesinlerin oldukça yüksek ve soğuk yerlerde yaşadığı düşünülüyor (genelde gece sıcaklığın 10 veya 5 °C'ye düşebildiği 1.000 ila 1.600 m yükseklikte). Bu yüzden daha alçak, daha sıcak yerlerde yaşayan erken insan türlerinin aksine sıcak kalmak için kıllara gereksinmiş olabilirler.[56] Yüksek enlemlerdeki H. erectus popülasyonlarında, D vitamini eksikliğini önlemek için daha açık ten rengi evrimleşmiş olabilir.[57] Türkiye'nin Denizli ilinde bulunan 300 ila 500 bin yıllık bir H. erectus örneğine, bilinen en eski tüberküloz menenjit vakası tanısı konuldu; bu vaka genellikle yüksek enlemlerde yaşayan koyu tenli insanlarda D vitamini eksikliği nedeniyle daha da kötüleşir.[58] Kılsızlaşmanın genelde terlemenin kolaylaşmasını sağladığı düşünülür,[59] ancak vücuttaki asalakların azaltılmasına yaradığı veya eşeysel seçilimle ilgili olduğu da önerilmiştir.[60][61]
Kültür
[değiştir | kaynağı değiştir]Toplumsal yapı
[değiştir | kaynağı değiştir]
Bir H. erectus topluluğuna ilişkin tek fosil kanıtı, Kenya'nın Ileret kentinin yakınlarındaki 4 bölgeden geliyor: burada bulunan 1,5 milyon yıllık 97 ayak izi muhtemelen en az 20 kişiden oluşan bir grup tarafından bırakılmıştı. Ayak izlerinin büyüklüğüne bakılırsa bu tamamen erkeklerden oluşan bir grup olabilir; bu da onların avcılık takımı, yiyecek arama takımı veya sınır devriyesi gibi özel bir görev grubu oldukları anlamına gelebilir. Bu eğer doğruysa, insan toplumlarını diğer büyük insansı maymunlardan ve toplumsal etobur memelilerden ayıran cinsiyete dayalı iş bölümünü de gösteriyor olacaktır. Büyük avları hedef alan modern avcı toplayıcı toplumlarda genellikle erkek gruplar yüksek riskli hayvanları avlamak için gönderilir ve düşük başarı oranı nedeniyle kadın gruplar daha öngörülebilir yiyeceklere odaklanır.[51] Günümüzün savanlarda yaşayan şempanze ve babun gruplarına dayanarak, H. erectus ergaster'in açık arazilerdeki büyük savan yırtıcılarına karşı savunma yapmak için büyük, çok erkekli gruplar hâlinde yaşadığı düşünülebilir.[62] Bununla birlikte H. erectus'un nüfus dağılımı modelleri, genellikle etobur yoğunluğunun yüksek olduğu alanlardan kaçındığını göstermektedir.[63] Boylardaki erkek-erkek bağı ve erkek-kadın arkadaşlıkları önemli toplumsal durumlar olabilir.[62]
H. erectus çocuklarının beyin büyüme hızları daha yüksek olduğundan, H. erectus muhtemelen modern insanlarla aynı düzeyde anne yatırımı veya çocuk yetiştirme davranışları göstermiyordu.[45]
H. erectus'un erkek ve dişilerinin diğer büyük insansı maymunlarla karşılaştırıldığında yaklaşık aynı büyüklükte olduğu düşünüldüğü için, genellikle tek eşli yaşadıkları varsayılır çünkü primatlarda eşeysel dimorfizmin azalması sıklıkla tek eşlilikle ilişkilendirilir.[48] Ancak H. erectus'ta eşeysel dimorfizmin modern insandakine benzer oranda olup olmadığı belirsizdir.[17] Eğer benzerse bu, ata türlere göre yalnızca dişilerin boyunun arttığı anlamına gelir. Dişi doğurganlığı veya beslenmesi değişmiş ve/veya erkekler üzerindeki büyük boyutlu olmaya yönelik baskı azalmış, sonuç olarak kadınların davranışlarında erkeklerin harem (tek bir baskın erkeğin liderliğindeki sürü) sürdürmesini zorlaştıracak bir değişim oluşmuştur.[64]
Beslenme
[değiştir | kaynağı değiştir]Beyin boyutunun artması genellikle daha etli bir beslenme ve bunun sonucunda daha yüksek kalori alımıyla doğrudan ilişkilendirilir. Protein tüketimindeki artışın H. erectus'un daha fazla böcek tüketmesinden kaynaklanabileceği de öne sürülmüştür. Ancak H. erectus'ta fazla enerji kullanan bağırsakların boyutunun küçülmüş olması da olanaklıdır. Çünkü büyük insansı maymunların bağırsağı, bitkisel maddeleri mayalayarak yağ sentezlemek için kullanılırdı, H. erectus'ta bunun yerini hayvansal yağlar alıyor ve enerji bağırsaklar yerine beyin büyümesi için kullanılıyor. Böylece H. erectus ata türlerle aynı kaloride besin tüketse de dolaylı olarak beyin boyutunu arttırmış olacaktır. H. erectus aynı zamanda et tüketimine bağımlılığın artışına bir tepki olarak avcı-toplayıcılık stratejisini kullanan ilk tür olabilir. Takım çalışmasına, işbölümüne ve yiyecek paylaşımına vurgu yapan avcı-toplayıcılık, önceki tarzlardan önemli ölçüde farklı bir geçim yoluydu.[40][65]

H. erectus'un yaşam alanları sıklıkla orta ila büyük boyutlu av hayvanları olan filler, gergedanlar, su aygırları, sığırlar ve yaban domuzu topluluklarıyla ilişkilendirilir. H. erectus büyük avlarla beslendiğinden, kayda değer miktarda yemek artığı olması gerekirdi. Avların etleri iyi değerlendirilmişse, gıda paylaşımı veya uzun süreli gıda saklama davranışının geliştiği varsayılabilir. H. erectus, boyutça büyümüş ve büyük avlara oldukça bağımlı olmuş olabilir. Buna kanıt olarak H. erectus'un Levant'ta ortadan kaybolması, Palaeoloxodon antiquus'un bölgesel olarak yok olmasıyla ilişkilendirilmiştir.[65] Bununla birlikte, H. erectus'un beslenmesi büyük olasılıkla konuma bağlı olarak değişiklik gösteriyordu. Örneğin, İsrail'deki 780 bin yıllık Gesher Benot Ya'aqov bölgesindeki H. erectus topluluğu 55 farklı türde meyve, sebze, tohum, kabuklu yemiş ve yumru kök toplayıp yiyordu, ayrıca başka türlü yenmeyecek olan bazı bitki bölümlerini kızartmak için ateşi kullanmışlardı. Fil ve alageyik gibi yaygın büyük hayvanların yanı sıra iki yaşamlılar, sürüngenler, kuşlar ve gerek sucul gerek karasal omurgasızlarla da beslendiler.[66] Kenya'nın Doğu Turkana Havzası'nda göl kenarında bulunan ve 1,95 milyon yıl öncesine tarihlenen FwJJ20 fosil bölgesindeki topluluk, (sığırlar, suaygırları ve gergedanların yanı sıra) kaplumbağa, timsah ve yayın balığı gibi sucul hayvanlarla besleniyordu. Büyük hayvanların leşleriyle beslenildiği düşünülse de kaplumbağalar ve balıklar avlanmıştı.[67] Doğu Afrika'da 2 ila 1,4 milyon yıl önce, sıcak mevsim otları ile beslenen toynaklıların leşleri, özellikle de Afrika antilopları (Alcelaphinae), H. erectus'un beslenmesinde giderek daha belirgin bir şekilde öne çıkıyordu.[68] Cava'da bulunan 1,5 milyon yıllık Trinil H. K. bölgesindeki H. erectus'un balık ve kabuklu deniz ürünleri ile beslendiği düşünülüyor.[69]
H. erectus'un dişleri ata türlerinki kadar çok yönlü değildi ve daha dar bir çeşitlilikteki yiyecekler için uygundu. Bununla birlikte aletler sert yiyecekleri ezmek veya parçalamak için kullanılmış, dolayısıyla beslenme çeşitliliğini eskisi gibi artırmış olabilir. Ayrıca bu yetenek, H. erectus'un farklı ortamlarda yaşamasına ve Afrika'nın ötesine göç etmesine olanak vermiş olabilir.[40]
1999'da İngiliz antropolog Richard Wrangham "yemek pişirme varsayımı"nı öne sürdü. Bu varsayım, H. erectus'un 2 milyon yıl önce ateş kullanma ve yemek pişirme yeteneğini kazanması ile atası H. habilis'ten ayrıldığını öne sürüyor. Böylece H. erectus'un beyin boyutunun 500 bin yıllık bir zaman diliminde H. habilis'tekinin iki katına çıkmasını ve özgün insan vücut planının ansızın ortaya çıkışını açıklıyor. Pişirme, protein sindirimini kolaylaştırır, besin emilimini hızlandırır ve besin kaynaklı patojenleri yok eder. Sonuç olarak topluluktaki birey sayısının artmasına olanak tanır, türde toplumsallaşma için seçilim baskısına neden olur ve daha fazla zihinsel beceri gerektirir.[70][71] Ancak fosil kayıtlarına göre H. erectus'un ortaya çıkışı ateş kullanımı ya da bu konuda herhangi bir teknolojik atılımla ilişkili gözükmüyor, muhtemelen yemek pişirme uygulaması 400 bin yıl öncesine kadar yaygın değildi.[40][65]
Cava insanının Güneydoğu Asya'ya yayılması, dev kaplumbağa Megalochelys'in bölgedeki yok oluşuyla çakışır. Kaplumbağanın epey bir süre saklanabilecek kolay bir av olduğu ve bu nedenle aşırı avlandığı sanılmaktadır.[72]
Teknoloji
[değiştir | kaynağı değiştir]Alet kullanımı
[değiştir | kaynağı değiştir]

H. erectus'un, Oldovan taş alet kültürünün yerini alan Aşölyen kültürünü icat ettiği kabul edilir.[73][74] Ayrıca 10 cm'den büyük taş yongaları ve el baltalarını (kazma, bıçak ve satır gibi yalnızca 2 yanı olan iki yüzeyli araçlar) ilk yapanlar da onlardı.[75] Bu el baltaları daha büyük ve daha ağırdı ancak daha keskin, yontulmuş uçları vardı.[76] Muhtemelen et, bitki veya odun kesmek gibi çeşitli etkinliklerde kullanılan çok amaçlı aletlerdi.[77] 1979'da Amerikalı paleontolog Thomas Wynn, Aşölyen teknolojisinin daha çok zeka kullanımı (öngörü ve planlama) gerektirdiğini ve standartlaşmamış biçim, kesit ve bakışım içeren Oldovan teknolojisinden belirgin şekilde daha karmaşık olduğunu belirtti. Buna dayanarak, H. erectus ile modern insan arasında zeka açısından önemli bir farklılık olmadığı ve son 300 bin yıldır zeka artışının kültürel evrim üzerinde önemli bir etken olmadığı sonucuna vardı.[78] Bununla birlikte, 1 yaşındaki bir H. erectus örneği, bu türün daha fazla beyin gelişimi için gereken uzun bir çocukluktan yoksun olduğunu ve dolayısıyla daha düşük bilişsel yeteneklerinin olduğunu gösteriyor.[79] Etiyopya'daki Melka Kunture, Kenya'daki Olorgesailie, Tanzanya'daki Isimila ve Zambiya'daki Kalambo Şelalesi gibi kimi bölgelerde muhtemelen birkaç nesil boyunca süren yerleşim nedeniyle toplu hâlde el baltaları bulunuyor.[77]
Aşölyen teknolojisinin en eski kalıntısı Kenya'nın Batı Turkana bölgesinde (1.76 myö) bulundu. Oldovan yontma taşları da bu bölgede bulunduğundan iki kültürün de bir süre birlikte var olduğu görülüyor. Aşölyen aletlerinin Afrika dışındaki en eski kalıntılarının 1 milyon yıllık olması, kültürün ancak Afrika'dan ikincil bir H. erectus göçünden sonra Avrasya'ya yayıldığını gösterir.[76]
Cava'daki Sangiran[80] ve Trinil'de[81] H. erectus'un hayvan kabuklarından alet ürettiğine dair kanıtlar bulundu. Çapı 6 ila 12 cm olan yuvarlak taşlar, Afrika ve Çin'in Alt Paleolitik kazı alanlarında sıklıkla bulunur. Bunların avcılıkta ipin ucuna bağlanarak kullanıldığı (bolas) düşünülüyor; bu eğer doğruysa, ip ve halat teknolojisinin H. erectus tarafından geliştirildiğini gösterir.[82]
Ateşin keşfi
[değiştir | kaynağı değiştir]H. erectus, ateşi kullanan ilk insan atası olarak anılır ancak ateşin ilk kez ne zaman kullanıldığı, kalıntılarının binlerce veya milyonlarca yıl gibi uzun zaman ölçeklerinde korunamaması nedeniyle tartışma konusudur. Öne sürülen en eski ateş yakılan bölgelerden ikisi de Kenya'da bulunan Koobi Fora'daki "FxJj20"[70][83][84] ve Chemoigut Oluşumu'ndaki "GnJi 1/6E" 1,5 milyon yıl önceye tarihlenirken[70][84] Güney Afrika'da bulunan Wonderwerk Mağarası'ndaki buluntular 1,7 milyon yıl öncesine tarihlenir.[85] İlk ateş kullanıcılarının mağaralarda yaşadığı ve doğal yangınların ürettiği ateşi uzun süre ya da fırsat ortaya çıktığında ara sıra kullandıkları düşünülüyor. Ateşi sürdürmek, ateş kullanan insanların tezek gibi yavaş yanan malzemeler konusunda bilgi sahibi olmasını gerektirirdi.[70] İnsan yapımı ateş kalıntıları, günümüzün 300 ila 400 bin yıl öncesinden sonra kazı alanlarında belirgin bir biçimde sıklaşır; bu, o dönemde ateş yakma tekniklerinde bazı ilerlemelerin gerçekleşmesi[70] ya da insan atalarının bu zamana dek ateşi yalnızca yangınlardan yararlanarak kullanması ile açıklanabilir.[40][65][84][86] Ateş yakma yöntemlerinin tek bir yerde bulunup tüm dünyaya yayılması yerine, farklı topluluklar tarafından bağımsız olarak birçok kez keşfedilip yitirilmesi ve yeniden keşfedilmesi mümkündür.[86] Ocaklara dair en eski izler, İsrail'deki Gesher Benot Ya'aqov'da bulunan en az 700 bin yıllık kalıntılardır. Bu ateş kalıntıları su kıyısında birden fazla katman hâlinde birikmiştir ve doğal yangınların ayırt edici özelliklerinden yoksundur.[71]
Yapay aydınlatma uyanıklık süresinin artmasını sağlamış olabilir (modern insanlar yaklaşık 16 saat uyanık kalır oysa diğer insansı maymunlar genellikle gün doğumundan gün batımına kadar uyanıktır) ve bu fazladan saatler toplumsallaşma için kullanılmış olabilir. Bu nedenle ateş kullanımının dilin kökeniyle de bağlantılı olduğu sanılır.[70][71] Ateş, yakındaki yırtıcıları uzakta tutarak insan atalarının ağaçlar yerine yerde uyumasını da mümkün kılmış olabilir.[71]
H. erectus'un buzul çağında soğuk iklimli Avrupa'ya göç edebilmesinin yalnız ateş kullanımı aracılığıyla mümkün olduğu düşünülür ancak Avrupa'da yaklaşık 300-400 bin yıl öncesine dek ateşin kullanıldığına dair kayda değer kanıt yoktur.[84] Eğer bu erken dönem Avrupalı H. erectus toplulukları ateş kullanmadıysa, nasıl sıcak kaldıkları, yırtıcılardan nasıl kaçındıkları, hayvansal yağ ve etleri nasıl tükettikleri büyük ölçüde belirsizdir. Daha kuzeydeki bölgelerde yıldırımın az görülmesi nedeniyle doğal yangınların da az sıklıkta olduğu düşünülür. Avrupalı H. erectus'un sadece belirli bölgelerde yangını nasıl sürdüreceğini biliyor olması ve yiyecekleri evden biraz uzakta pişirmiş olması da mümkündür; eğer öyleyse, ateş izleri ile H. erectus yerleşimlerinin izleri birbirinden çok uzaktadır.[71] Belki de H. erectus yalnızca ılık iklimli buzullar arası dönemlerde kuzeye doğru göç etmiş olabilir (bu nedenle ateşe, yiyecek saklamaya veya giysiye ihtiyaç duymamıştır),[87] dağılım modelleri onların genellikle daha sıcak alt-orta enlemlerde yaşadığını göstermektedir.[63] Kuzey Çin'deki Zhoukoudian mağarasında 770 bin yıl önce görece soğuk bir iklimde yaşamış Pekin insanının, ateş yakma becerisinin olup olmadığı belirsizdir.[88]
Giyim
[değiştir | kaynağı değiştir]_Fundort_Nariokotome,_Kenia,_Rekonstruktion_im_Neanderthal_Museum.jpg)
Giysilerin ne zaman icat edildiği büyük ölçüde belirsizdir. En erken tahminlere göre giysiler, ısı yalıtan vücut kıllarının eksikliğini kapatmak için 3 milyon yıl kadar önce kullanılmaya başlanmıştır.[55] Modern insanlarlarda yaşayan saç ve vücut bitlerinin atalarının (ikincisi yalnızca giysili bireylerde yaşayabilir) yaklaşık 170 bin yıl önce, modern insanlar Afrika'dan göç etmeden çok önce, birbirinden ayrıldığı biliniyor. Bu, insanların soğuk iklimlerde yaşamadan önce de giysi kullandığı anlamına gelir. Hayvan derisinin ilk olarak giyim amacıyla kullanıldığı düşünülür. Deri yüzmeyle ilişkilendirilen en eski kalıntılar yaklaşık 780 bin yıl öncesine tarihlenir ancak bunların giyim için kullanıldığı üzerine bir kanıt yoktur.[89]
Dilin evrimi
[değiştir | kaynağı değiştir]1984 yılında, 1,6 milyon yıl önce yaşamış ergenlik çağındaki bir birey olan Turkana Çocuğu'nun omurgası incelendi ve konuşmayı sağlayacak solunum kaslarının gerektiği gibi gelişmediği ortaya atıldı. 2001 yılında Amerikalı insan bilimciler Bruce Latimer ve James Ohman, Turkana Çocuğu'nun iskelet displazisi (olağan dışı büyüme) ve skolyoz rahatsızlıklarının olduğu sonucuna vardı.[90] 2006 yılında Amerikalı insan bilimci Marc Meyer ve meslektaşları, 1,8 milyon yıllık H. e. georgicus örneğinin omurga biçiminin modern insanlarda görülen çeşitlilik aralığı içinde olduğunu öne sürerek Turkana Çocuğu'nun omurga stenozunun olduğunu ve türün temsilcilerinden biri olmadığını savladı. Ayrıca Meyer, H. e. georgicus'un Afrikalı olmayan tüm H. erectus'ların atası olduğunu varsayarak H. e. ergaster dışındaki tüm H. erectus'larda solunum kaslarının türü ses çıkarma veya konuşmadan alıkoymadığı sonucuna vardı.[91] Ancak insan bilimci Regula Schiess ve meslektaşları, 2013 ve 2014 yıllarında yaptıkları çalışmalarda Turkana Çocuğu'nda herhangi bir doğuştan kusur üstüne bir kanıt bulamadı ve örneğin türün temsilcisi olduğunu belirledi.[92][93]
Sinirsel olarak tüm insan türlerinin beyinleri benzer yapıdadır ve aynı şekilde H. erectus'un modern insanlarda cümle kurma ve konuşmadan sorumlu olan Broca ve Wernicke alanları modern insanlarla karşılaştırılabilir niteliktedir. Ancak bu, yetişkin şempanzelerde bile büyük Broca alanlarına rastlandığından, konuşma yeteneği açısından herhangi bir şeyin göstergesi değildir. Ayrıca bu bölgelerin arkaik insanlarda dil için kullanıp kullanılmadığı belirsizdir.[94] 1 yaşındaki bir H. erectus örneği, bireylerin dilin kazanılacağı beyin gelişimi için yeteri kadar uzun bir çocukluk döneminden geçmediğini göstermektedir.[79]
Dil kemiği, dili destekler ve sesin perdesini kontrol etmek için ses yolunun değişimini mümkün kılar. İtalya'nın Castel di Guido kasabasında bulunan ve 400 bin yıl öncesine tarihlenen bir H. erectus dil kemiği, Australopithecus'takinden çok kendisinden sonraki insan türlerindekine benzer olarak çubuk şeklindedir. Bununla birlikte kemik, kas izlerinden yoksundur ve kalkan biçiminde bir yapıya sahiptir; ayrıca dil kemiğindeki cornu majus modern insanlardakine göre küçüktür. Bu, H. erectus'un modern insan gibi konuşması için uygun bir ses aygıtının olmadığı anlamına gelir.[95] Teknolojik gelişmeyle birlikte beyin boyutunun ve kültürel karmaşıklığın artışı ve Neandertaller ile modern insanların konuşma yeteneklerini son ortak atadan miras aldığı varsayımı üzerinden H. erectus'un biraz ön-dil kullandığı ve daha sonra ortaya çıkacak gelişmiş dilllerin temel yapısını oluşturduğu çıkarılabilir.[96] Ancak konuşma yeteneğine sahip son ortak ata H. heidelbergensis de olabilir. İspanya'daki Sima de los Huesos Mağarası'nda bulunan 530 bin yıllık bir H. heidelbergensis örneğinin dil kemiği, modern insandakine benzerdir,[97] bölgeden alınan başka bir örneğin ise insan konuşmasını algılayabilecek kadar hassas bir işitsel yetisinin olduğu düşünülüyor.[98]
Soyunun tükenmesi
[değiştir | kaynağı değiştir]2019 yılında yayımlanan bir çalışmaya göre son H. erectus topluluğu 117 ila 108 bin yıl önce Cava Adası'ndaki Ngandong'da yaşamıştır.[5]
2020'de araştırmacılar, H. erectus ve H. heidelbergensis'in, soyları tükenmeden hemen önce, iklimine uyum sağladıkları bölgelerin yarısından fazlasını kaybettiğini, yayılım alanlarında ise buna karşılık gelen bir azalma olmadığını ve iklim değişikliğinin, geçmişteki insan türlerinin yok olmasında önemli bir etken olduğunu belirledi.[99][100][101]
Fosiller
[değiştir | kaynağı değiştir]
Çin'deki Zhoukoudian Mağarası'nın alt mağarası dünya çapındaki en önemli kazı alanlarından biridir.[102] Mağarada 45 ayrı H. erectus'a ait olan kalıntıların yanı sıra binlerce taş alet bulundu.[102] 1951'de Çin'de yeniden keşfedilen iki kafatası parçası ve "Ejder Kemiği Tepesi"nde bulunan dört diş dışındaki buluntular 2. Dünya Savaşı sırasında kayboldu.[102]
2016 yılında araştırmacılar, Homo erectus'un daha önce düşünüldüğü gibi benzersiz derecede kalın kafatası kemiklerine sahip olmadığını belirledi.[103]
Bireysel fosiller
[değiştir | kaynağı değiştir]Başlıca H. erectus fosilleri:
- Endonezya (Cava Adası): Trinil 2 (holotip), Sangiran koleksiyonu, Sambungmachan koleksiyonu[104] ve Ngandong koleksiyonu
- Çin ("Pekin insanı"): Lantiyen (Gongwangling ve Chenjiawo), Yunxian, Zhoukoudian, Nanjing, Hexian
- Kenya: KNM ER 3883 ve KNM ER 3733
- Vietnam: Kuzey, Tham Khuyen,[105] Hoa Binh[kaynak belirtilmeli]
- Gürcistan: Dmanisi koleksiyonu ("Homo erectus georgicus")
- Etiyopya: Daka kafatası
- Eritre: Buia kafatası (olası bir H. ergaster)[106]
- Denizli, Türkiye: Kocabaş kafatası[58]
- Drimolen, Güney Afrika: DNH 134[107]
Soy oluş
[değiştir | kaynağı değiştir]Aşağıda, Homo cinsinin, soyların ne zaman ayrıldığını da gösteren, 2021'de yayımlanmış kladogramı bulunmaktadır.[108]
| Homo (2.85) |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
H. erectus ilk olarak Afrika'da ortaya çıktı. Homo heidelbergensis (soyu Homo sapiens'i de içerir), Asyalı H. erectus'tan evrimleşti. Aynı zaman aralıklarında yaşayan türlerin birbirleriyle çiftleştiği üzerine kanıtlar vardır; bu nedenle yukarıdaki gibi herhangi bir soy oluş, insan türlerininin evrimi hakkında yalnızca kabataslak bir izlenim verir. Soyu tükenmiş türler, diğerleriyle melezleşerek kısmen varlığını sürdürmüş olabilir. Yukarıdaki zaman aralıklarında yaşamış olan insansılar Homo floresiensis, Homo naledi, Homo luzonensis, Homo rudolfensis, Australopithecus sediba, Australopithecus africanus türleri ve Paranthropus cinsi dâhil edilmemiştir.
Resim galerisi
[değiştir | kaynağı değiştir]-
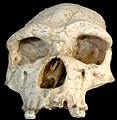 Homo erectus tautavelensis kafatası
Homo erectus tautavelensis kafatası -
 Tautavel, Fransa'da bulunan Homo erectus'un alt çenesinin kopyası
Tautavel, Fransa'da bulunan Homo erectus'un alt çenesinin kopyası -
 Özgün "Sangiran II" kafatası, Koenigswald koleksiyonu, Senckenberg Müzesi
Özgün "Sangiran II" kafatası, Koenigswald koleksiyonu, Senckenberg Müzesi -
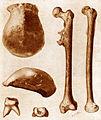 Pithecanthropus erectus'un (şimdiki Homo erectus) ilk fosilleri 1891'de Cava'da bulundu.
Pithecanthropus erectus'un (şimdiki Homo erectus) ilk fosilleri 1891'de Cava'da bulundu. -
 Etiyopya'da bulunan Daka kafatasından elde edilen kanıtlara dayanan bir yeniden canlandırma
Etiyopya'da bulunan Daka kafatasından elde edilen kanıtlara dayanan bir yeniden canlandırma


.jpg)


Ayrıca bakınız
[değiştir | kaynağı değiştir]- Kozarnika
- Fosil alanları listesi (bağlantı dizini ile birlikte)
- İnsan evriminin fosil listesi (resimlerle birlikte)
Kaynakça
[değiştir | kaynağı değiştir]- ^ a b Herries, Andy I. R.; Martin, Jesse M.; Leece, A. B.; Adams, Justin W.; Boschian, Giovanni; Joannes-Boyau, Renaud; Edwards, Tara R.; Mallett, Tom; Massey, Jason; Murszewski, Ashleigh; Neubauer, Simon (3 Nisan 2020). "Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa". Science (İngilizce). 368 (6486). doi:10.1126/science.aaw7293. ISSN 0036-8075. 23 Eylül 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ekim 2021.
- ^ Dembo, Mana; Radovčić, Davorka; Garvin, Heather M.; Laird, Myra F.; Schroeder, Lauren; Scott, Jill E.; Brophy, Juliet; Ackermann, Rebecca R.; Musiba, Chares M.; de Ruiter, Darryl J.; Mooers, Arne Ø. (Ağustos 2016). "The evolutionary relationships and age of Homo naledi: An assessment using dated Bayesian phylogenetic methods". Journal of Human Evolution (İngilizce). 97: 17-26. doi:10.1016/j.jhevol.2016.04.008. 20 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ van den Bergh, Gerrit D.; Kaifu, Yousuke; Kurniawan, Iwan; Kono, Reiko T.; Brumm, Adam; Setiyabudi, Erick; Aziz, Fachroel; Morwood, Michael J. (Haziran 2016). "Homo floresiensis-like fossils from the early Middle Pleistocene of Flores". Nature (İngilizce). 534 (7606): 245-248. doi:10.1038/nature17999. ISSN 1476-4687. 23 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Détroit, Florent; Mijares, Armand Salvador; Corny, Julien; Daver, Guillaume; Zanolli, Clément; Dizon, Eusebio; Robles, Emil; Grün, Rainer; Piper, Philip J. (Nisan 2019). "A new species of Homo from the Late Pleistocene of the Philippines". Nature (İngilizce). 568 (7751): 181-186. doi:10.1038/s41586-019-1067-9. ISSN 1476-4687. 4 Ocak 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ a b Rizal, Yan; Westaway, Kira E.; Zaim, Yahdi; van den Bergh, Gerrit D.; Bettis, E. Arthur; Morwood, Michael J.; Huffman, O. Frank; Grün, Rainer; Joannes-Boyau, Renaud; Bailey, Richard M.; Sidarto (Ocak 2020). "Last appearance of Homo erectus at Ngandong, Java, 117,000–108,000 years ago". Nature (İngilizce). 577 (7790): 381-385. doi:10.1038/s41586-019-1863-2. ISSN 1476-4687. 19 Ekim 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Mart 2022.
- ^ Ben‐Dor, Miki; Sirtoli, Raphael; Barkai, Ran (Ağustos 2021). "The evolution of the human trophic level during the Pleistocene". American Journal of Physical Anthropology (İngilizce). 175 (S72): 27-56. doi:10.1002/ajpa.24247. ISSN 0002-9483. 12 Şubat 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Zohar, Irit; Alperson-Afil, Nira; Goren-Inbar, Naama; Prévost, Marion; Tütken, Thomas; Sisma-Ventura, Guy; Hershkovitz, Israel; Najorka, Jens (14 Kasım 2022). "Evidence for the cooking of fish 780,000 years ago at Gesher Benot Ya'aqov, Israel". Nature Ecology & Evolution (İngilizce). 6 (12): 2016-2028. doi:10.1038/s41559-022-01910-z. ISSN 2397-334X. PMID 36376603. 5 Ekim 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ a b Theunissen 2012.
- ^ Yen, Hsiao-pei (1 Kasım 2014). "Evolutionary Asiacentrism, Peking Man, and the Origins of␣Sinocentric Ethno-Nationalism". Journal of the History of Biology (İngilizce). 47 (4): 585-625. doi:10.1007/s10739-014-9381-4. ISSN 1573-0387.
- ^ a b c Sigmon 1981.
- ^ "Zhoukoudian: Geography and Culture". Encyclopedia of Global Archaeology. Springer Science+Business Media. 2014. ss. 7961-7965. doi:10.1007/978-1-4419-0465-2_1899. ISBN 978-1-4419-0466-9.
- ^ "Java Man". Encyclopedia of Global Archaeology. 2014. ss. 4182-4188. doi:10.1007/978-1-4419-0465-2_712. ISBN 978-1-4419-0426-3.
- ^ Darwin, Charles (1871). The Descent of Man and Selection in Relation to Sex. John Murray. ISBN 978-0-8014-2085-6.
- ^ van Wyhe, John; Kjærgaard, Peter C. (1 Haziran 2015). "Going the whole orang: Darwin, Wallace and the natural history of orangutans". Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences (İngilizce). 51: 53-63. doi:10.1016/j.shpsc.2015.02.006. ISSN 1369-8486. PMID 25861859.
- ^ Curnoe, D. (Haziran 2010). "A review of early Homo in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (Homo gautengensis sp. nov.)". HOMO. 61 (3): 151-177. doi:10.1016/j.jchb.2010.04.002. ISSN 0018-442X.
- ^ de Lumley, Marie-Antoinette (1 Haziran 2015). "L'homme de Tautavel. Un Homo erectus européen évolué. Homo erectus tautavelensis". L'Anthropologie. Paléoanthropologie. L'Homme de Tautavel. 119 (3): 303-348. doi:10.1016/j.anthro.2015.06.001. ISSN 0003-5521. 25 Ekim 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ a b c Spoor, F.; Leakey, M. G.; Gathogo, P. N.; Brown, F. H.; Antón, S. C.; McDougall, I.; Kiarie, C.; Manthi, F. K.; Leakey, L. N. (Ağustos 2007). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya". Nature (İngilizce). 448 (7154): 688-691. doi:10.1038/nature05986. ISSN 1476-4687. 17 Mayıs 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Zhu, Zhaoyu; Dennell, Robin; Huang, Weiwen; Wu, Yi; Qiu, Shifan; Yang, Shixia; Rao, Zhiguo; Hou, Yamei; Xie, Jiubing; Han, Jiangwei; Ouyang, Tingping (Temmuz 2018). "Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago". Nature (İngilizce). 559 (7715): 608-612. doi:10.1038/s41586-018-0299-4. ISSN 1476-4687. 25 Eylül 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ "Tools from China are oldest hint of human lineage outside Africa". Nature. 2018. doi:10.1038/d41586-018-05696-8. ISSN 0028-0836. 17 Ağustos 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Li, Hao; Li, ChaoRong; Kuman, Kathleen (1 Mart 2017). "Longgudong, an Early Pleistocene site in Jianshi, South China, with stratigraphic association of human teeth and lithics". Science China Earth Sciences (İngilizce). 60 (3): 452-462. doi:10.1007/s11430-016-0181-1. ISSN 1869-1897.
- ^ "Our direct human ancestor Homo erectus is older than we thought". EurekAlert. AAAS. 3 Nisan 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, ve diğerleri. (June 2011). "Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma". Proceedings of the National Academy of Sciences of the United States of America. 108 (26). ss. 10432-10436. Bibcode:2011PNAS..10810432F. doi:10.1073/pnas.1106638108. PMC 3127884 $2. PMID 21646521.
- ^ Agustí, Jordi; Lordkipanidze, David (Haziran 2011). "How "African" was the early human dispersal out of Africa?". Quaternary Science Reviews (İngilizce). 30 (11-12): 1338-1342. doi:10.1016/j.quascirev.2010.04.012. 16 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Rightmire, G. Philip (1998). "Human evolution in the Middle Pleistocene: The role ofHomo heidelbergensis". Evolutionary Anthropology: Issues, News, and Reviews (İngilizce). 6 (6): 218-227. doi:10.1002/(SICI)1520-6505(1998)6:6<218::AID-EVAN4>3.0.CO;2-6.
- ^ Asfaw, Berhane; Gilbert, W. Henry; Beyene, Yonas; Hart, William K.; Renne, Paul R.; WoldeGabriel, Giday; Vrba, Elisabeth S.; White, Tim D. (Mart 2002). "Remains of Homo erectus from Bouri, Middle Awash, Ethiopia". Nature (İngilizce). 416 (6878): 317-320. doi:10.1038/416317a. ISSN 0028-0836. 9 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Husson, Laurent; Salles, Tristan; Lebatard, Anne-Elisabeth; Zerathe, Swann; Braucher, Régis; Noerwidi, Sofwan; Aribowo, Sonny; Mallard, Claire; Carcaillet, Julien; Natawidjaja, Danny H.; Bourlès (8 Kasım 2022). "Javanese Homo erectus on the move in SE Asia circa 1.8 Ma". Scientific Reports (İngilizce). 12 (1): 19012. doi:10.1038/s41598-022-23206-9. ISSN 2045-2322. PMC 9643487 $2. PMID 36347897.
- ^ Zaim, Yahdi; Ciochon, Russell L.; Polanski, Joshua M.; Grine, Frederick E.; Bettis, E. Arthur; Rizal, Yan; Franciscus, Robert G.; Larick, Roy R.; Heizler, Matthew; Aswan; Eaves, K. Lindsay (Ekim 2011). "New 1.5 million-year-old Homo erectus maxilla from Sangiran (Central Java, Indonesia)". Journal of Human Evolution (İngilizce). 61 (4): 363-376. doi:10.1016/j.jhevol.2011.04.009. 9 Aralık 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ There was long-standing uncertainty whether H. floresiensis should be considered close to H. erectus, close to H. sapiens, or an altogether separate species. In 2017, it was suggested on morphological grounds that H. floresiensis is a sister species to either H. habilis or to a minimally habilis-erectus-ergaster-sapiens clade, and its line much more ancient than Homo erectus itself. "The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters". Journal of Human Evolution. 107: 107-133. June 2017. doi:10.1016/j.jhevol.2017.02.006. PMID 28438318.
- ^ Kennedy, Kenneth A. R.; Sonakia, Arun; Chiment, John; Verma, K. K. (Aralık 1991). "Is the Narmada hominid an Indian Homo erectus ?". American Journal of Physical Anthropology (İngilizce). 86 (4): 475-496. doi:10.1002/ajpa.1330860404. ISSN 0002-9483. 2 Ekim 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Kaifu, Yousuke; Baba, Hisao; Aziz, Fachroel; Indriati, Etty; Schrenk, Friedemann; Jacob, Teuku (Aralık 2005). "Taxonomic affinities and evolutionary history of the early Pleistocene hominids of Java: Dentognathic evidence". American Journal of Physical Anthropology (İngilizce). 128 (4): 709-726. doi:10.1002/ajpa.10425. ISSN 0002-9483. 9 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Krantz, G.S. (1975). "An explanation for the diastema of Javan erectus Skull IV". In: Paleoanthropology, Morphology and Paleoecology. La Hague: Mouton, 361–372.
- ^ "Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia" (PDF). Nature Ecology & Evolution. 3 (5): 755-764. May 2019. doi:10.1038/s41559-019-0860-z. PMID 30962558. 25 Aralık 2023 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 5 Mayıs 2024.
- ^ Baba H, Aziz F, Kaifu Y, Suwa G, Kono RT, Jacob T (February 2003). "Homo erectus calvarium from the Pleistocene of Java". Science. 299 (5611). ss. 1384-1388. doi:10.1126/science.1081676. PMID 12610302.
- ^ a b Balzeau A (2006). "Are thickened cranial bones and equal participation of the three structural bone layers autapomorphic traits of Homo erectus?". Bulletins et mémoires de la Société d'Anthropologie de Paris. 18 (3–4). ss. 145-163. doi:10.4000/bmsap.1528. 30 Mart 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Copes LE, Kimbel WH (January 2016). "Cranial vault thickness in primates: Homo erectus does not have uniquely thick vault bones". Journal of Human Evolution. Cilt 90. ss. 120-134. doi:10.1016/j.jhevol.2015.08.008. PMID 26767964.
- ^ Franciscus RG, Trinkaus E (April 1988). "Nasal morphology and the emergence of Homo erectus". American Journal of Physical Anthropology. 75 (4). ss. 517-527. doi:10.1002/ajpa.1330750409. PMID 3133950.
- ^ Jacobs LF (February 2019). "The navigational nose: a new hypothesis for the function of the human external pyramid". The Journal of Experimental Biology. 222 (Pt Suppl 1). s. jeb186924. doi:10.1242/jeb.186924. PMID 30728230.
- ^ a b Antón, Susan C.; Taboada, Hannah G.; Middleton, Emily R.; Rainwater, Christopher W.; Taylor, Andrea B.; Turner, Trudy R.; Turnquist, Jean E.; Weinstein, Karen J.; Williams, Scott A. (5 Temmuz 2016). "Morphological variation in Homo erectus and the origins of developmental plasticity". Philosophical Transactions of the Royal Society B: Biological Sciences (İngilizce). 371 (1698): 20150236. doi:10.1098/rstb.2015.0236. ISSN 0962-8436. 9 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Clark, Gary; Henneberg, Maciej (2022). "Interpopulational variation in human brain size: Implications for hominin cognitive phylogeny". Anthropological Review. 84 (4): 405-429. doi:10.2478/anre-2021-0029.
- ^ a b c d e Ungar, Peter S.; Grine, Frederick E.; Teaford, Mark F. (1 Ekim 2006). "Diet in Early Homo : A Review of the Evidence and a New Model of Adaptive Versatility". Annual Review of Anthropology (İngilizce). 35 (1): 209-228. doi:10.1146/annurev.anthro.35.081705.123153. ISSN 0084-6570. 25 Ekim 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Mart 2022.
- ^ Alan Walker, Richard Leakey (1993). The Nariokotome Homo erectus skeleton. Harvard University Press. s. 412. ISBN 9780674600751. 1 Mayıs 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Ekim 2022.
- ^ "The Effects of Mortality, Subsistence, and Ecology on Human Adult Height and Implications for Homo Evolution". Current Anthropology. 53 (S6): 359-368. 2012. doi:10.1086/667694.
- ^ Antón, Susan C.; Taboada, Hannah G.; Middleton, Emily R.; Rainwater, Christopher W.; Taylor, Andrea B.; Turner, Trudy R.; Turnquist, Jean E.; Weinstein, Karen J.; Williams, Scott A. (5 Temmuz 2016). "Morphological variation in Homo erectus and the origins of developmental plasticity". Philosophical Transactions of the Royal Society B: Biological Sciences. 371 (1698): 20150236. doi:10.1098/rstb.2015.0236. ISSN 0962-8436. PMC 4920293 $2. PMID 27298467.
- ^ Antón, Susan C.; Potts, Richard; Aiello, Leslie C. (4 Temmuz 2014). "Evolution of early Homo: An integrated biological perspective". Science (İngilizce). 345 (6192): 1236828. doi:10.1126/science.1236828. ISSN 0036-8075. PMID 24994657. 24 Mart 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ a b Simpson, Scott W.; Quade, Jay; Levin, Naomi E.; Butler, Robert; Dupont-Nivet, Guillaume; Everett, Melanie; Semaw, Sileshi (14 Kasım 2008). "A Female Homo erectus Pelvis from Gona, Ethiopia". Science (İngilizce). 322 (5904): 1089-1092. doi:10.1126/science.1163592. ISSN 0036-8075. 16 Mart 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ a b Leakey, Mary D (1979). Olduvai Gorge: my search for early man (İngilizce). Londra: Collins. ISBN 9780002116138. OCLC 647137093.
- ^ Rightmire G. Philip (1990). The Evolution of Homo Erectus: Comparative Anatomical Studies of an Extinct Human Species. Cambridge University Press. 9 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ a b Plavcan, J. Michael (Aralık 2012). "Body Size, Size Variation, and Sexual Size Dimorphism in Early Homo". Current Anthropology (İngilizce). 53 (S6): S409-S423. doi:10.1086/667605. ISSN 0011-3204. 27 Aralık 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Ruff C (March 2008). "Femoral/humeral strength in early African Homo erectus". Journal of Human Evolution. 54 (3). ss. 383-390. doi:10.1016/j.jhevol.2007.09.001. PMID 17977577.
- ^ Antón, Susan C.; Taboada, Hannah G.; Middleton, Emily R.; Rainwater, Christopher W.; Taylor, Andrea B.; Turner, Trudy R.; Turnquist, Jean E.; Weinstein, Karen J.; Williams, Scott A. (5 Temmuz 2016). "Morphological variation in Homo erectus and the origins of developmental plasticity". Philosophical Transactions of the Royal Society B: Biological Sciences. 371 (1698). s. 20150236. doi:10.1098/rstb.2015.0236. ISSN 0962-8436. PMC 4920293 $2. PMID 27298467.
- ^ a b Hatala KG, Roach NT, Ostrofsky KR, Wunderlich RE, Dingwall HL, Villmoare BA, ve diğerleri. (July 2016). "Footprints reveal direct evidence of group behavior and locomotion in Homo erectus". Scientific Reports. 6 (28766). s. 28766. Bibcode:2016NatSR...628766H. doi:10.1038/srep28766. PMC 4941528 $2. PMID 27403790.
- ^ Roach, & Richmond. (2015). "Clavicle length, throwing performance and the reconstruction of the Homo erectus shoulder". Journal of Human Evolution, 80(C), 107–113.
- ^ Haeusler M, Schiess R, Boeni T (November 2011). "New vertebral and rib material point to modern bauplan of the Nariokotome Homo erectus skeleton" (PDF). Journal of Human Evolution. 61 (5). ss. 575-582. doi:10.1016/j.jhevol.2011.07.004. PMID 21868059. 9 Mart 2024 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 5 Mayıs 2024.
- ^ Rogers AR, Iltis D, Wooding S (2004). "Genetic Variation at the MC1R Locus and the Time since Loss of Human Body Hair". Current Anthropology. 45 (1). ss. 105-108. doi:10.1086/381006.
- ^ a b Gilligan I (2010). "The Prehistoric Development of Clothing: Archaeological Implications of a Thermal Model". Journal of Archaeological Method and Theory. Cilt 15. ss. 15-80. doi:10.1007/s10816-009-9076-x.
- ^ Dávid-Barrett T, Dunbar RI (May 2016). "Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling". Journal of Human Evolution. Cilt 94. ss. 72-82. doi:10.1016/j.jhevol.2016.02.006. PMC 4874949 $2. PMID 27178459.
- ^ Jablonski NG (March 2012). "Human skin pigmentation as an example of adaptive evolution". Proceedings of the American Philosophical Society. 156 (1). ss. 45-57. JSTOR 23558077. PMID 23035389.
- ^ a b Kappelman J, Alçiçek MC, Kazanci N, Schultz M, Ozkul M, Sen S (January 2008). "First Homo erectus from Turkey and implications for migrations into temperate Eurasia". American Journal of Physical Anthropology. 135 (1). ss. 110-116. doi:10.1002/ajpa.20739. PMID 18067194.
- ^ Best A, Kamilar JM (April 2018). "The evolution of eccrine sweat glands in human and nonhuman primates". Journal of Human Evolution. Cilt 117. ss. 33-43. doi:10.1016/j.jhevol.2017.12.003. PMID 29544622.
- ^ Pagel M, Bodmer W (2004). "The Evolution of Human Hairlessness: Cultural Adaptations and the Ectoparasite Hypothesis". Evolutionary Theory and Processes: Modern Horizons. Springer, Dordrecht. ss. 329-335. doi:10.1007/978-94-017-0443-4_17. ISBN 978-94-017-0443-4.
- ^ Gile J (2010). "Naked Love: The Evolution of Human Hairlessness". Biological Theory. 5 (4). ss. 326-336. doi:10.1162/BIOT_a_00062.
- ^ a b Willems EP, van Schaik CP (August 2017). "The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates". Journal of Human Evolution. Cilt 109. ss. 11-21. doi:10.1016/j.jhevol.2017.05.003. PMID 28688456.
- ^ a b Carotenuto F, Tsikaridze N, Rook L, Lordkipanidze D, Longo L, Condemi S, Raia P (June 2016). "Venturing out safely: The biogeography of Homo erectus dispersal out of Africa". Journal of Human Evolution. Cilt 95. ss. 1-12. doi:10.1016/j.jhevol.2016.02.005. hdl:10356/82274. PMID 27260171.
- ^ Plavcan JM (2012). "Implications of Male and Female Contributions to Sexual Size Dimorphism for Inferring Behavior in the Hominin Fossil Record". International Journal of Primatology. 33 (6). ss. 1364-1381. doi:10.1007/s10764-012-9642-z.
- ^ a b c d Ben-Dor M, Gopher A, Hershkovitz I, Barkai R (2011). "Man the fat hunter: the demise of Homo erectus and the emergence of a new hominin lineage in the Middle Pleistocene (ca. 400 kyr) Levant". PLOS ONE. 6 (12). ss. e28689. Bibcode:2011PLoSO...628689B. doi:10.1371/journal.pone.0028689. PMC 3235142 $2. PMID 22174868.
- ^ Melamed Y, Kislev ME, Geffen E, Lev-Yadun S, Goren-Inbar N (December 2016). "The plant component of an Acheulian diet at Gesher Benot Ya'aqov, Israel". Proceedings of the National Academy of Sciences of the United States of America. 113 (51). ss. 14674-14679. Bibcode:2016PNAS..11314674M. doi:10.1073/pnas.1607872113. PMC 5187744 $2. PMID 27930293.
- ^ Steele TE (June 2010). "A unique hominin menu dated to 1.95 million years ago". Proceedings of the National Academy of Sciences of the United States of America. 107 (24). ss. 10771-10772. Bibcode:2010PNAS..10710771S. doi:10.1073/pnas.1005992107. PMC 2890732 $2. PMID 20534542.
- ^ Patterson, D. B.; Braun, D. R.; Behrensmeyer, A. K.; Merritt, S.; Zliobaite, I.; Reeves, J. S.; Wood, B. A.; Fortelius, M.; Bobe, R. (1 Eylül 2017). "Ecosystem evolution and hominin paleobiology at East Turkana, northern Kenya between 2.0 and 1.4 Ma". Palaeogeography, Palaeoclimatology, Palaeoecology. Cilt 481. ss. 1-13. Bibcode:2017PPP...481....1P. doi:10.1016/j.palaeo.2017.05.001.
- ^ Joordens JC, Wesselingh FP, de Vos J, Vonhof HB, Kroon D (December 2009). "Relevance of aquatic environments for hominins: a case study from Trinil (Java, Indonesia)". Journal of Human Evolution. 57 (6). ss. 656-671. doi:10.1016/j.jhevol.2009.06.003. PMID 19683789.
- ^ a b c d e f Gowlett JA (June 2016). "The discovery of fire by humans: a long and convoluted process". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371 (1696). s. 20150164. doi:10.1098/rstb.2015.0164. PMC 4874402 $2. PMID 27216521.
- ^ a b c d e Gowlett JA, Wrangham RW (2013). "Earliest fire in Africa: Towards the convergence of archaeological evidence and the cooking hypothesis". Azania: Archaeological Research in Africa. 48 (1). ss. 5-30. doi:10.1080/0067270X.2012.756754.
- ^ Rhodin A, Pritchard P, van Dijk PP, Saumure R, Buhlmann K, Iverson J, Mittermeier R, (Ed.) (2015). Conservation Biology of Freshwater Turtles and Tortoises. First. 5. Chelonian Research Foundation. s. 15. doi:10.3854/crm.5.000e.fossil.checklist.v1.2015. ISBN 978-0-9653540-9-7. 10 Ağustos 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Beck RB, Black L, Krieger LS, Naylor PC, Shabaka DI (1999). World History: Patterns of Interaction. Evanston, IL: McDougal Littell. ISBN 978-0-395-87274-1.[sayfa belirt]
- ^ Richards MP (December 2002). "A brief review of the archaeological evidence for Palaeolithic and Neolithic subsistence". European Journal of Clinical Nutrition. 56 (12). ss. 1270-1278. doi:10.1038/sj.ejcn.1601646. PMID 12494313.
- ^ de la Torre I (July 2016). "The origins of the Acheulean: past and present perspectives on a major transition in human evolution". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371 (1698). s. 20150245. doi:10.1098/rstb.2015.0245. PMC 4920301 $2. PMID 27298475.
- ^ a b Lepre CJ, Roche H, Kent DV, Harmand S, Quinn RL, Brugal JP, ve diğerleri. (August 2011). "An earlier origin for the Acheulian". Nature. 477 (7362). ss. 82-85. Bibcode:2011Natur.477...82L. doi:10.1038/nature10372. PMID 21886161.
- ^ a b Nowell A, Chang ML (2009). "The Case Against Sexual Selection as an Explanation of Handaxe Morphology" (PDF). PaleoAnthropology. ss. 77-88. 2 Ağustos 2020 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 5 Mayıs 2024.
- ^ Wynn T (1979). "The Intelligence of Later Acheulean Hominids". Man. 14 (3). ss. 371-391. doi:10.2307/2801865. JSTOR 2801865.
- ^ a b Coqueugniot H, Hublin JJ, Veillon F, Houët F, Jacob T (September 2004). "Early brain growth in Homo erectus and implications for cognitive ability". Nature. 431 (7006). ss. 299-302. Bibcode:2004Natur.431..299C. doi:10.1038/nature02852. PMID 15372030.
- ^ Choi K, Driwantoro D (2007). "Shell tool use by early members of Homo erectus in Sangiran, central Java, Indonesia: cut mark evidence". Journal of Archaeological Science. 34 (1). ss. 48-58. Bibcode:2007JArSc..34...48C. doi:10.1016/j.jas.2006.03.013.
- ^ Joordens JC, d'Errico F, Wesselingh FP, Munro S, de Vos J, Wallinga J, ve diğerleri. (February 2015). "Homo erectus at Trinil on Java used shells for tool production and engraving". Nature. 518 (7538). ss. 228-231. Bibcode:2015Natur.518..228J. doi:10.1038/nature13962. PMID 25470048.
- ^ Turner J (1996). History and Science of Knots. World Scientific. ss. 6-8. ISBN 9789810224691. 13 Mayıs 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mayıs 2024.
- ^ Hlubik S, Berna F, Feibel C, Braun D (2017). "Researching the Nature of Fire at 1.5 Mya on the Site of FxJj20 AB, Koobi Fora, Kenya, Using High-Resolution Spatial Analysis and FTIR Spectrometry". Current Anthropology. Cilt 58. ss. S243-S257. doi:10.1086/692530.
- ^ a b c d Roebroeks W, Villa P (March 2011). "On the earliest evidence for habitual use of fire in Europe". Proceedings of the National Academy of Sciences of the United States of America. 108 (13). ss. 5209-5214. Bibcode:2011PNAS..108.5209R. doi:10.1073/pnas.1018116108. PMC 3069174 $2. PMID 21402905.
- ^ Beaumont PB (2011). "The Edge: More on Fire-Making by about 1.7 Million Years Ago at Wonderwerk Cave in South Africa". Current Anthropology. 52 (4). ss. 585-595. doi:10.1086/660919.
- ^ a b Sandgathe D (2017). "Identifying and Describing Pattern and Process in the Evolution of Hominin Use of Fire". Current Anthropology. Cilt 58. ss. S360-S370. doi:10.1086/691459. hdl:11858/00-001M-0000-002C-0141-3.
- ^ Antón SC (2003). "Natural history of Homo erectus". American Journal of Physical Anthropology. 122 (S37). ss. 126-170. doi:10.1002/ajpa.10399. PMID 14666536.
- ^ Zhong M, Shi C, Gao X, Wu X, Chen F, Zhang S, Zhang X, Olsen JW (2013). "On the possible use of fire by Homo erectus at Zhoukoudian, China". Chinese Science Bulletin. 59 (3). ss. 335-343. doi:10.1007/s11434-013-0061-0.
- ^ Toups MA, Kitchen A, Light JE, Reed DL (January 2011). "Origin of clothing lice indicates early clothing use by anatomically modern humans in Africa". Molecular Biology and Evolution. 28 (1). ss. 29-32. doi:10.1093/molbev/msq234. PMC 3002236 $2. PMID 20823373.
- ^ Latimer B, Ohman J (2001). "Axial dysplasia in Homo erectus". Journal of Human Evolution. Cilt 40.
- ^ Meyer M, Lordkipanidze D, Vekua A (2006). Language and empathy in Homo erectus: Behaviors suggested by a modern spinal cord from Dmanisi, but not Nariokotome. Annual meeting of the Paleoanthroplogy Society. San Juan, Puerto Rico.
- ^ Schiess R, Haeusler M (March 2013). "No skeletal dysplasia in the Nariokotome boy KNM-WT 15000 (Homo erectus)--a reassessment of congenital pathologies of the vertebral column". American Journal of Physical Anthropology. 150 (3). ss. 365-374. doi:10.1002/ajpa.22211. PMID 23283736.
- ^ Schiess R, Boeni T, Rühli F, Haeusler M (February 2014). "Revisiting scoliosis in the KNM-WT 15000 Homo erectus skeleton" (PDF). Journal of Human Evolution. 67 (48–59). ss. 48-59. doi:10.1016/j.jhevol.2013.12.009. PMID 24491377. 10 Şubat 2024 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 5 Mayıs 2024.
- ^ Luef EM (2018). "Tracing the human brain's classical language areas in extant and extinct hominids". The talking species: Perspectives on the evolutionary, neuronal and cultural foundations of language. Uni-Press Graz. ISBN 978-3-902666-52-9.
- ^ Capasso L, Michetti E, D'Anastasio R (December 2008). "A Homo erectus hyoid bone: possible implications for the origin of the human capability for speech". Collegium Antropologicum. 32 (4). ss. 1007-1011. PMID 19149203.
- ^ Hillert DG (2015). "On the Evolving Biology of Language". Frontiers in Psychology. Cilt 6. s. 1796. doi:10.3389/fpsyg.2015.01796. PMC 4656830 $2. PMID 26635694.
- ^ Martínez I, Arsuaga JL, Quam R, Carretero JM, Gracia A, Rodríguez L (January 2008). "Human hyoid bones from the middle Pleistocene site of the Sima de los Huesos (Sierra de Atapuerca, Spain)" (PDF). Journal of Human Evolution. 54 (1). ss. 118-124. doi:10.1016/j.jhevol.2007.07.006. PMID 17804038. 19 Ekim 2020 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 5 Mayıs 2024.
- ^ Martínez I, Rosa M, Arsuaga JL, Jarabo P, Quam R, Lorenzo C, ve diğerleri. (July 2004). "Auditory capacities in Middle Pleistocene humans from the Sierra de Atapuerca in Spain". Proceedings of the National Academy of Sciences of the United States of America. 101 (27). ss. 9976-9981. Bibcode:2004PNAS..101.9976M. doi:10.1073/pnas.0403595101. PMC 454200 $2. PMID 15213327.
- ^ Padmanaban D (6 Kasım 2020). "Climate Change May Have Been a Major Driver of Ancient Hominin Extinctions". SAPIENS. 9 Kasım 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Kasım 2020.
- ^ "Climate change likely drove early human species to extinction, modeling study suggests". phys.org (İngilizce). 12 Kasım 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Kasım 2020.
- ^ Raia P, Mondanaro A, Melchionna M, Di Febbraro M, Diniz-Filho JA, Rangel TF, ve diğerleri. (23 Ekim 2020). "Past Extinctions of Homo Species Coincided with Increased Vulnerability to Climatic Change". One Earth (İngilizce). 3 (4). ss. 480-490. Bibcode:2020OEart...3..480R. doi:10.1016/j.oneear.2020.09.007. hdl:2158/1211341. ISSN 2590-3330.
- ^ a b c Zanolli, Clément, et al. "Inner Tooth Morphology of Homo Erectus from Zhoukoudian. New Evidence from an Old Collection Housed at Uppsala University, Sweden." Journal of Human Evolution, vol. 116, Mar. 2018, pp. 1–13.
- ^ Copes, Lynn E., and William H. Kimbel. "Cranial Vault Thickness in Primates: Homo Erectus Does Not Have Uniquely Thick Vault Bones." Journal of Human Evolution, vol. 90, Jan. 2016, pp. 120–134.
- ^ Delson E, Harvati K, Reddy D, Marcus LF, Mowbray K, Sawyer GJ, ve diğerleri. (Nisan 2001). "The Sambungmacan 3 Homo erectus calvaria: a comparative morphometric and morphological analysis". The Anatomical Record. Cilt 262. ss. 380-397. doi:10.1002/ar.1048. PMID 11275970.
- ^ Ciochon R, Long VT, Larick R, González L, Grün R, de Vos J, ve diğerleri. (April 1996). "Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam". Proceedings of the National Academy of Sciences of the United States of America. 93 (7). ss. 3016-3020. Bibcode:1996PNAS...93.3016C. doi:10.1073/pnas.93.7.3016. PMC 39753 $2. PMID 8610161.
- ^ Schuster AM (September–October 1998). "New Skull from Eritrea". Archaeology. 1 Ekim 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Ekim 2015.
- ^ Herries AI, Martin JM, Leece AB, Adams JW, Boschian G, Joannes-Boyau R, ve diğerleri. (April 2020). "Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa". Science. 368 (6486). ss. eaaw7293. doi:10.1126/science.aaw7293. hdl:11568/1040368. PMID 32241925.
- ^ Ni, Xijun; Ji, Qiang; Wu, Wensheng; Shao, Qingfeng; Ji, Yannan; Zhang, Chi; Liang, Lei; Ge, Junyi; Guo, Zhen; Li, Jinhua; Li, Qiang; Grün, Rainer; Stringer, Chris (28 Ağustos 2021). "Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage". The Innovation (İngilizce). 2 (3). s. 100130. Bibcode:2021Innov...200130N. doi:10.1016/j.xinn.2021.100130. ISSN 2666-6758. PMC 8454562 $2. PMID 34557770.
Konuyla ilgili yayınlar
[değiştir | kaynağı değiştir]- Leakey R, Walker A (November 1985). "Homo Erectus Unearthed". National Geographic. 168 (5). ss. 624-629. ISSN 0027-9358. OCLC 643483454.
- Sigmon BA, Cybulski JS (1981). Homo erectus: Papers in Honor of Davidson Black. University of Toronto Press. JSTOR 10.3138/j.ctvcj2jdw.11.
- Theunissen B, Theunissen LT (2012). Eugène Dubois and the Ape-Man from Java. Springer Netherlands. ISBN 9789400922099.
Dış bağlantılar
[değiştir | kaynağı değiştir] Vikitür'de Homo erectus ile ilgili ayrıntılı taksonomik bilgiler bulunur.
Vikitür'de Homo erectus ile ilgili ayrıntılı taksonomik bilgiler bulunur. Wikimedia Commons'ta Homo erectus ile ilgili çoklu ortam belgeleri bulunur
Wikimedia Commons'ta Homo erectus ile ilgili çoklu ortam belgeleri bulunur- Bradshaw Foundation'da Homo erectus 11 Şubat 2021 tarihinde Wayback Machine sitesinde arşivlendi.
- Arkeolojiinfo.com 16 Mayıs 2011 tarihinde Wayback Machine sitesinde arşivlendi.'da Homo erectus
- Homo erectus 5 Haziran 2021 tarihinde Wayback Machine sitesinde arşivlendi. humanorigins.si.edu
- Possible co-existence with Homo Habilis 1 Mart 2009 tarihinde Wayback Machine sitesinde arşivlendi. – BBC News
- John Hawks Kocabaş fosili 15 Nisan 2021 tarihinde Wayback Machine sitesinde arşivlendi.
- Homo erectus
- The Age of Homo erectus 16 Nisan 2016 tarihinde Wayback Machine sitesinde arşivlendi. – Homo erectus'un seyahatinin interaktif haritası
- Human Timeline (Interactive) 17 Temmuz 2017 tarihinde Wayback Machine sitesinde arşivlendi. – Smithsonian, National Museum of Natural History (Ağustos 2016).
- Homo erectus
- Geçiş fosilleri
- 1894'te tanımlanan fosil taksonları
- Erken insan türleri
- Buzul Çağı primatları
- Pliosen devri primatları
- Tarih öncesi Anadolu
- Tarih öncesi Çin
- Tarih öncesi Etiyopya
- Tarih öncesi Gürcistan
- Tarih öncesi Macaristan
- Tarih öncesi Hindistan
- Tarih öncesi Endonezya
- Tarih öncesi Kenya
- Tarih öncesi İspanya
- Tarih öncesi Tanzanya