
外側膝状体
| 脳: 外側膝状体 | |
|---|---|
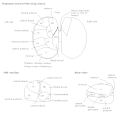
 後脳と中脳;後外側から見た図(外側膝状体は一番上にマークされている) | |
| 名称 | |
| 日本語 | 外側膝状体 |
| 英語 | lateral geniculate nucleus |
| ラテン語 | nucleus geniculatus lateralis |
| 略号 | LGN, LGB, LG |
| 関連構造 | |
| 上位構造 | 視床 |
| 動脈 | 前脈絡叢動脈、後脈絡叢動脈 |
| 静脈 | 視床線状体静脈 |
| 画像 | |
| アナトモグラフィー | 三次元CG |
| Digital Anatomist |
視床 視放線 冠状断(黒質) 水平断(膝状体) 視放線 |
| 関連情報 | |
| Brede Database | 階層関係、座標情報 |
| NeuroNames | 関連情報一覧 |
| NIF | 総合検索 |
| MeSH | Geniculate+Bodies |
| グレイ解剖学 | 書籍中の説明(英語) |
外側膝状体(がいそくしつじょうたい, lateral geniculate nucleus,LGN,lateral geniculate body,LGB)は、脳の視床領域の一部であり、中枢神経系の網膜から情報を受け取り、視覚情報の処理を行う。
LGNは網膜から視神経、視交叉、視索を通って直接情報を受け取る。一次視覚野に視放線を通して直接投射する。また、LGNには一次視覚野からのフィードバック入力も多く投射している。
網膜神経節細胞が軸索を伸ばし、視神経としてLGNに投射している。
構造
[編集]左右のLGNは固有の層構造を持つ。"geniculate"(膝状)とは"膝のように折れ曲がった"との意味。ヒトやマカクザルなど多くの霊長類では、LGNの各層のあいだに神経網(neruopil)があり、LGNニューロンの各層が"ケーキ"、神経網がアイシングとなって互いをはさみこむ、サンドイッチ状ないしはケーキ状の構造となっている。ヒトやマカクザルでは、LGNは大細胞層(magnocellular layer)が2層、小細胞層(parvocellular layer)が4層の、6層構造を持つとされる。こうした層の数は、霊長類の種によって異なり、種によっては別の層が加わることもある。
M, P, K 細胞
[編集]| 種類 | 大きさ | 機能 | 位置 |
| M: 大細胞(Magnocellular cells) | 大きな細胞体 | 情報処理に要する時間は短い。高速な処理ができるが、詳細な処理はできない | 1層と2層 |
| P: 小細胞(Parvocellular cells, "parvicellular"とも) | 小さい細胞体 | 情報処理に要する時間は長い。低速な処理であるが、詳細な処理が可能である。たとえば、P細胞は色の情報を処理できる。M細胞はこうした処理はおこなわない。 | 3, 4, 5層と6層 |
| K: 顆粒細胞(Koniocellular cells, "interlaminar"とも) | 非常に小さな細胞体 | M細胞やP細胞ほどは広く知られていない。 層のあいだに位置する。K細胞は機能的・神経化学的にM細胞やP細胞とは異なり、視覚野への第三のチャンネルとなっている。K細胞系が視知覚において果たす役割は、現在のところよく分かっていない。しかし、視覚と体性感覚・固有感覚情報の連合や、色覚への関与などが提案されている。 | M層とP層の層間 |

大細胞、小細胞、顆粒細胞の層は、類似する名称の神経節細胞と対応している。
小細胞層と大細胞層の神経線維は、以前はUngerleider-Mishkinの腹側系と背側系に対応すると考えられていた。しかし、近年の研究では、2つの処理経路は両者の神経線維をともに含んでいることが示されている[1]。
他の重要な網膜-皮質経路として網膜視蓋路がある。これは、上丘や視床枕核を経て、後頭頂葉や内側側頭葉へ至る経路である。
同側と反対側
[編集]M細胞とP細胞の分類に加えて、層は以下のように分類される:[2]
- LGNと同側(ipsilateral)の目は第2, 3, 5層に情報を送る
- LGNと反対側(contralateral)の目は第1, 4, 6層に情報を送る。
英��での簡単な記憶法は、"See I? I see, I see"を覚えることである。ここで、"see"は"contralateral"のCを表し、"I"は"ipsilateral"のIを表す。
こうした記述は、多くの霊長類のLGNに当てはまるが、全てではない。同側と反対側の目からこのように情報を受け取る方法は、メガネザルでは異なっている[3]。神経科学者には、このように一見するとメガネザルと他の霊長類に差があるように見えることが、メガネザルが比較的昔に生じた種であり、霊長類の進化とは独立だったことを示す、と考えるものもある。[4]。
視覚において、右側の目は右視野だけでなく、左視野の情報も受け取ることは、注意するべきである。このことは、左目を閉じれば分かる:右目によって、視野の右側も左側も見えるだろう。ただし、左視野の一部は、鼻によって遮蔽されている。
LGNでは、左右の目で対応する視野位置の情報は"積み重なって"おり、クラブサンドイッチの第1層から6層までトゥースピックで貫いたとすると、同じ視野位置を6回貫通することになる。
LGNへの入力
[編集]LGNは網膜からの入力を受ける。
いくつかの生物種では、LGNは視蓋(哺乳類での上丘)からの入力も受ける[5]。
LGNの出力
[編集]LGNから出力される情報は視放線として、内包のレンズ核後部を形成する。
LGNを出た軸索は、V1へと向かう。大細胞層1-2層と小細胞層3-6層は、ともにV1の4層にシナプスを作り、4cβは小細胞層から、4cαは大細胞層から入力を受ける。しかし、顆粒細胞層(1-6層の層間)は、V1の第2,3層へ軸索を送る。V1の6層からはLGNへフィードバックする軸索が送られる。
盲視の研究により、LGNからの神経投射はV1だけではなく高次の視覚領野にも及ぶことが示唆されている。盲視の患者はある視野位置での知覚を行うことができないが、こうした患者にテストを行うと、視野内の情報が無意識的にコードされていることが示される。このことは、LGNからの神経線維が、一次視覚野と高次視覚野の双方に投射することを示すと考えられることがある。
視覚における機能
[編集]LGNの機能はよく分かっていない。網膜は中心周辺拮抗処理により空間的無相関化(spatial decorrelation)を行うのに対して、LGNは時間的無相関化を行うと考えられることがある。こうした時空間無相関化により、より効率的なコーディングが可能になる。しかし、それ以上の処理が行われていることは確実だと考えられている。
視床の他の領域、特に他の中継核(relay nuclei)と同じように、LGNは視覚系が注意の焦点を特定の重要な情報に向けるのに役立つとされる。このことは、体の少し左側で音が聞こえたとすると、聴覚系が視覚系に対して、視覚的注意を空間のその領域に向けるよう、LGNを通じて"伝える"というものである。
LGNでは受容野がより精細化される場所でもある。
近年のヒトfMRI研究によると、空間的注意とサッカード眼球運動の双方がLGNの活動を修飾するとされる。
他の画像
[編集]-
 視床
視床 -
 脳幹の解剖。側面から見た図
脳幹の解剖。側面から見た図 -
 視神経と視索の走行を示す模式図
視神経と視索の走行を示す模式図 -
 視床核
視床核 -
 視索の3D模式図
視索の3D模式図





脚注
[編集]- ^ Goodale & Milner, 1993, 1995.
- ^ Nicholls J., et. al. From Neuron to Brain: Fourth Edition. Sinauer Associates, Inc. 2001.
- ^ Rosa MG, Pettigrew JD, Cooper HM (1996) Unusual pattern of retinogeniculate projections in the controversial primate Tarsius. Brain Behav Evol 48(3):121-129.
- ^ Collins CE, Hendrickson A, Kaas JH (2005) Overview of the visual system of Tarsius. Anat Rec A Discov Mol Cell Evol Biol 287(1):1013-1025.
- ^ In Chapter 7, section "The Parcellation Hypothesis" of "Principals of Brain Evolution", Georg Striedter (Sinauer Associates, Sunderland, MA, USA, 2005)によると"...we now know that the LGN receives at least some inputs from the optic tectum (or superior colliculus) in many amniotes". 著者は"Wild, J.M. 1989. Pretectal and tectal projections to the homolog of the dorsal lateral geniculate nucleus in the pigeon - an anterograde and retrograde tracing study with cholera-toxin conjugated to horseradish-peroxidase. Brain Res 489: 130-137" および "Kaas, J.H., and Huerta, M.F. 1988. The subcortical visual system of primates. In: Steklis H. D., Erwin J., editors. Comparative primate biology, vol 4: neurosciences. New York: Alan Liss, pp. 327-391."を引用している。
外部リンク(英語)
[編集]- Blohm G and Schreiber C. LGN in the visual pathway. Retrieved September 1, 2004.
- Malpeli J. Malpeli Lab Home Page. Retrieved September 1, 2004.